

- Home
Welcome !
Welcome to john houghton's home page for his biology courses. This site is designed as a hub for curating and sharing lectures, course syllabi, assignments, and links to relevant resources. Use the menu bar at the top of the screen to navigate through the site.
(Please note: this page is currently under construction.)

- BIOL 2107
Fall '22 CRN87989
Lectures: (1)
- Courses

Lecture 13: Extensions of Mendelian Genetics
![]()
![]()
pdf download video with sound... Click "here" or watch below.
|
Extensions of Mendelian Genetics
Incomplete dominance example?
Codominance example?
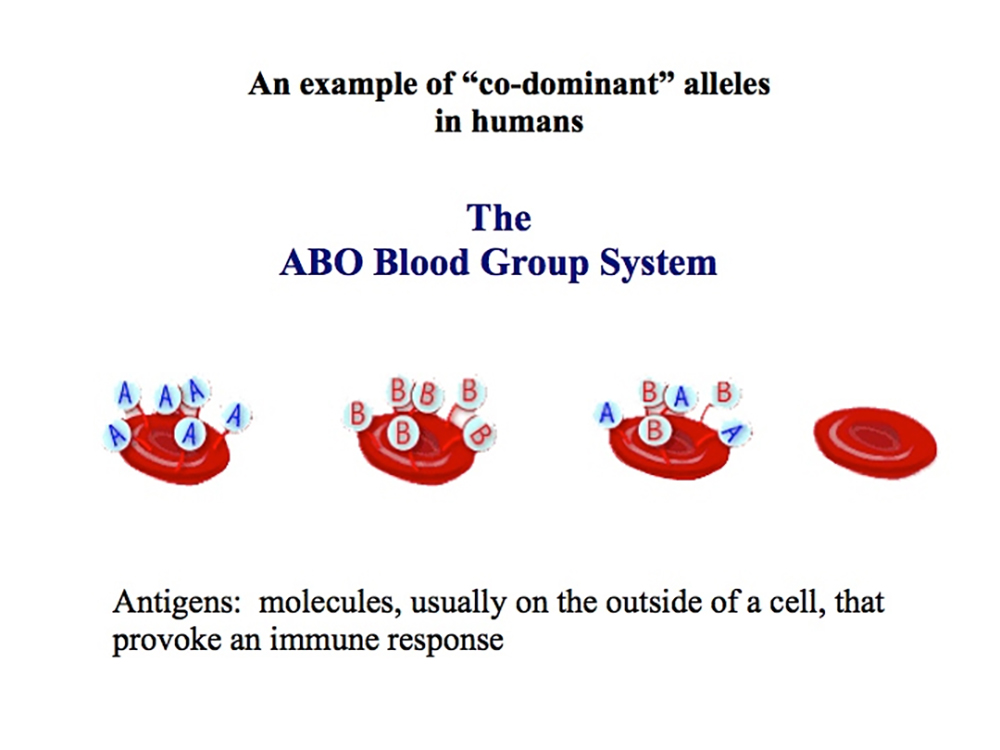
Multiple Alleles:The "ABO" blood-type system
The ABO system is now known to be a polymorphism of complex carbohydrate structures comprised of glycoproteins and glycolipids expressed at the surface of red blood cells.
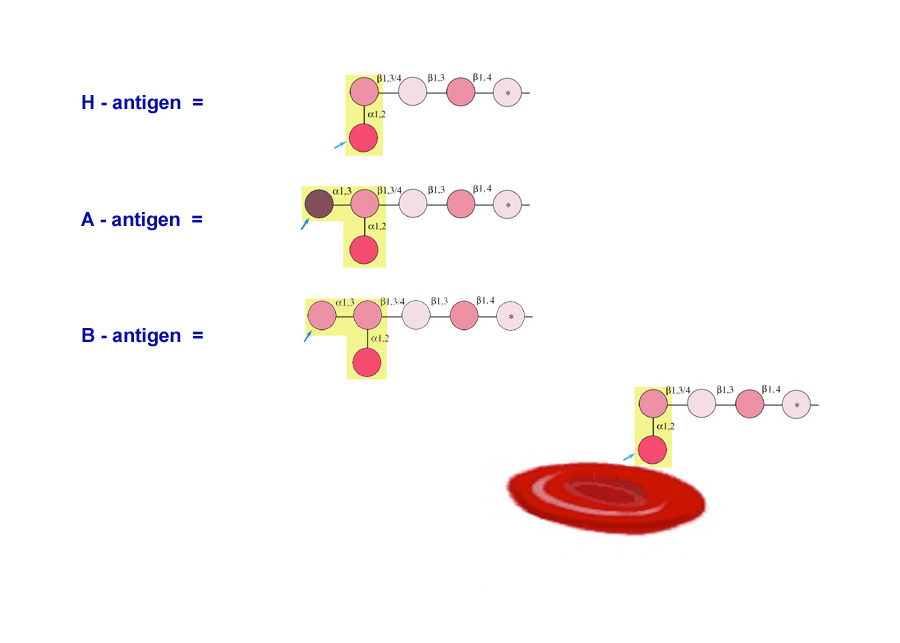
The alleles for blood type are IA, IBand IO. They all occupy one locus.
These alleles determine which antigens (proteins) are present on the surface
of red blood cells.
These "antigens" react with proteins called antibodies in the serum of certain individuals and cause the blood to clot.

The "ABO" blood-type system, however, is not just a series of antigens on the surface of each of red blood cell, but also feature their corresponding "antibodies" that exist within the blood serum.
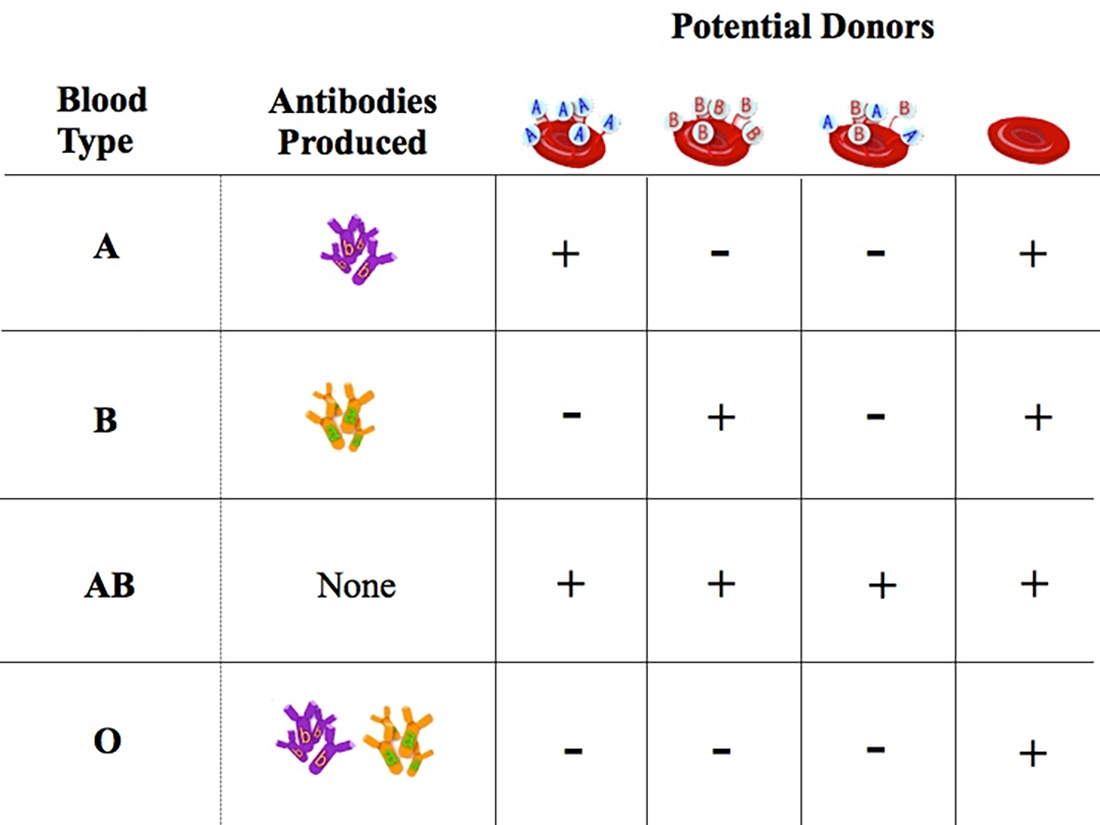
If A antigen (blood cell) and A antibody (serum) are present in the blood... the blood will clot!!.
Therefore, the result of mixing blood from each of the different ABO blood types can result in red blood cell agglutination, or clumping, which may prove to be fatal.
| Blood Type | Genotypes | ABO
Enzymes Present |
RBC
Antigens Present |
Serum Antibodies |
|---|---|---|---|---|
"A" |
IA IA,
IA IO |
"H", "A" | A, H | anti-B |
"B" |
IB IB,
IB IO |
"H", "B" | B, H | anti-A |
"AB" |
IA IO |
"H", "A", "B" | A, B, H | none |
"O" |
IO IO |
"H" | H | anti-A,
anti-B |
Chart Of A-B-O Blood Donor & Recipient Compatibility
|
Note that blood-types A and B can be considered to be "codominant", as the presence of both alleles AB gives rise to a completely different phenotype, when compared to a homozgous AA or a BB equivalent.
What about Rhesus factors?... THIS IS NOT REQUIRED these are a whole new class of antigens, discovered when blood from "rhesus monkeys" was injected into guinea pigas (circa 1940's). There are over 50 different types of Rh factors, but the most comomonly known one is the D antigen (Rho[D]), which -if it is present- indicates that that person is Rh-positive; if the D antigen is absent, that person is Rh-negative.
In contrast to the ABO system, however, antibodies to Rh antigens aren't necesssarily inherent to the person's blood, and can develop as an immune response after a blood transfusion, or during pregnancy.
At a mother's first antenatal screening, blood tests are taken in order to determine her blood type (A, B, AB or O) as well as her rhesus status (Rh-positive or Rh-negative).
If the mother has the rhesus factor (which is a protein on the surface of her red blood cells) then she is said to be Rh-positive. If not, then she is Rh-negative. 85% of people are Rh-positive.
The rhesus state only really begins to play a role during pregnancy if the mother is Rh-negative, the father is Rh-positive and the baby is also Rh-positive.
Rh(D) positive cells contain the D antigen, which can stimulate Rh(d) negative blood to produce harmful antibodies that can destroy red cells. The harmful antibody is called ‘anti-D’ antibody, and can be produced by a mother who is Rh-negative carrying a baby who is Rh-positive.
Rhesus incompatibility doesn’t cause any problems with a first pregnancy because (unlike the AB antibodies) the rhesus antibodies aren’t inherently present in the mother’s blood.
However, in subsequent pregnancies, if the babies are Rh-positive, there may be a problem. The mother’s antibodies can / will cross over the placenta into the baby’s blood causing a reaction.
This causes problems with the baby’s haemoglobin levels (the iron-carrying element in the red blood cells) which could then fall, causing anaemia. Blood transfusion would then be necessary (see chart below) at birth and the babies could also be severely jaundiced.
Gene Interactions
Some phenotypes are the result of the expression of more than one gene. As such, expression of these different genes can give rise to gene products that alter the effects (phenotypes) of these other genes.
Epistasis occurs when the alleles of one gene cover up or alter the expression of alleles of another gene.
"the interaction of genes that are not alleles, in particular the suppression of the effect of one such gene by another."
Given that genes of any organism do not operate in total isolation from one another, but obviously are functioning in a common cellular environment. a phenomenon that occurs when the alleles of one gene covers up (or alters) the expression of alleles of another gene.


Labrador coat colour is an example of Epistasis in that there are two genes which affect coat colour -the "E" gene, which affects the presence of a dark pigment in the coat and the "B" gene, which governs the degree to which that pigment is present in the coat, where B_ gives rise to a "black lab", but only if the genotypes at the E locus are E_ or EE.
While blood groups would also be a more than adequate example of this phenomenon, a quite overt example of epistatic effect, albeit more indirect, would be life and death consequences!
Several Genes interacting
In a similar way to fur colour in rabbits another example of genes interacting is fur coat colour in mice in the example shown in the picture:
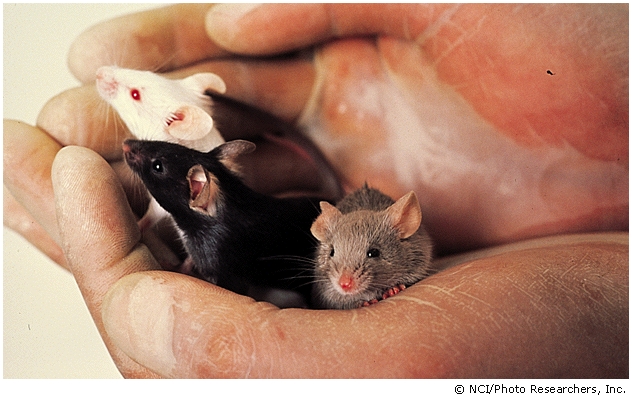
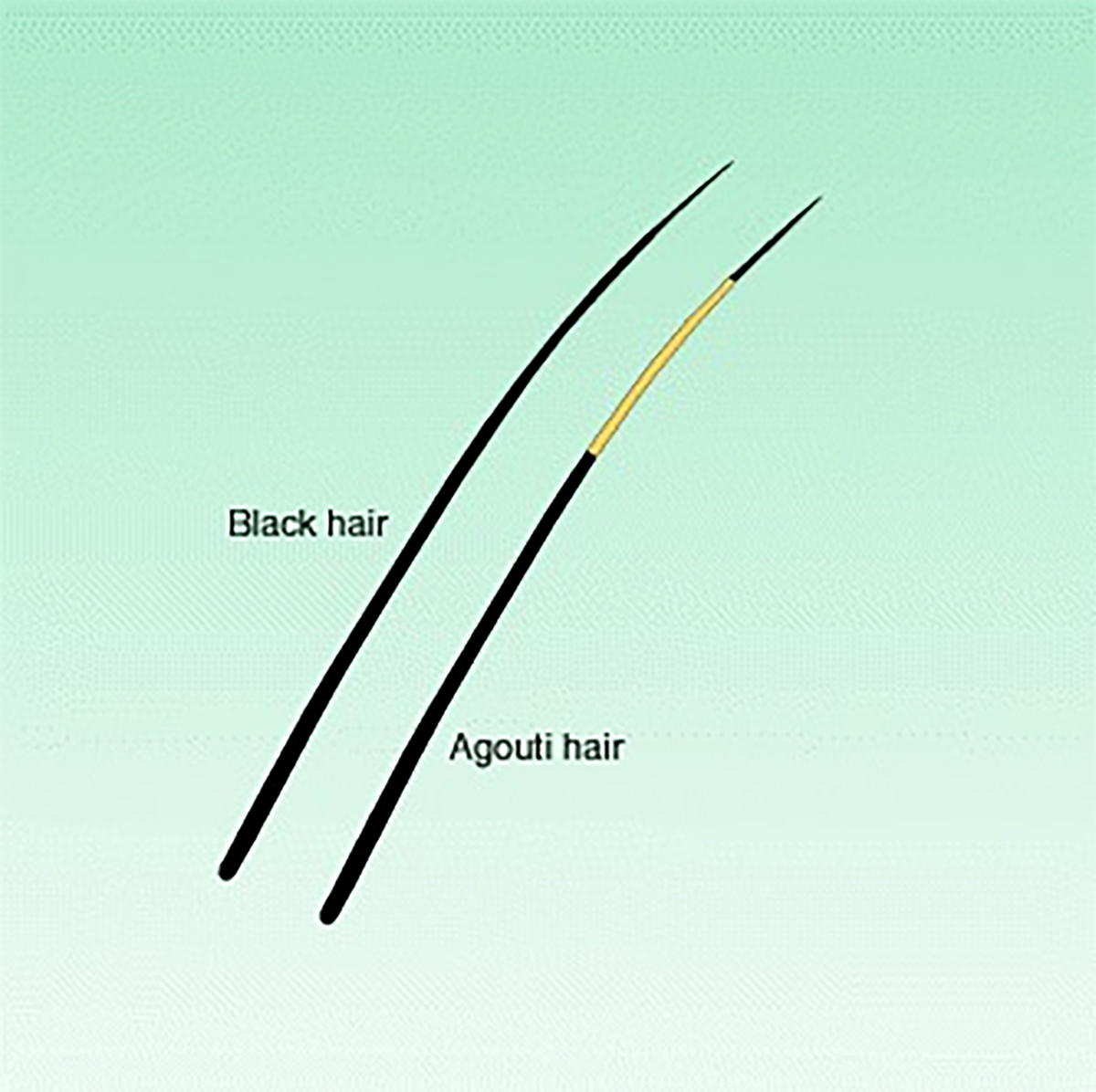
The A allele determines a banded pattern, called agouti.
The recessive a allele results in unbanded hairs.
The genotypes AA or Aa are, therefore, agouti.The genotype BB results in fur colour that is solidly black, whereas bb denotes brown fur colour .
Another gene, at the C-locus -an entirely different locus- determines if any coloration occurs at all.
The genotypes CC and Cc have colour, whereas the double recessive cc are albino, which do not allow any colour to show through as the cc genotype blocks all pigment production.
Mice that are heterozygous for ALL THREE GENES genes are agouti.
The F2 phenotypic ratio of fur colour in an initial parental cross between a BB, AA and a bb, aa would be: 9 agouti: 3 cinnamon: 3 Black and 1 Brown:The corresponding genotypes are 9 agouti (1 BBAA + 2 BbAA + 4 BbAa): 3 cinnamon (1 bbAA + 2 bbAa): 3 black (1 BBaa + 2 Bbaa) and 1 brown (1 bbaa).
In addition there are other genes... D and S which control "dilution" of these traits as well as "distribution -throughout the body".. (patches or spots)".
Lethal Genes

In this instance the same gene that affects coat colour in the mouse also has some influence upon development of the embryo. Thus, one gene, does not always mean one function. If, as in this case, It has more than one function and is therefore considerred to be pleiotropic
Other examples of gene/allelic expression, which differ from expected patterns of Medelian inheritance are:
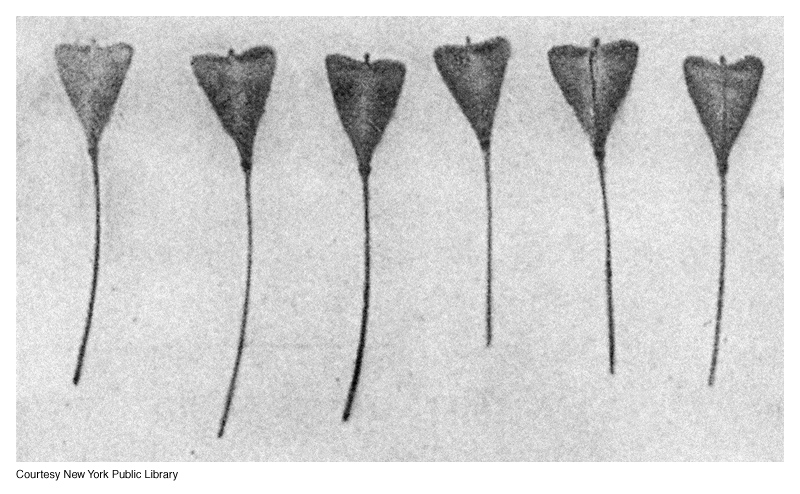
Duplicate genes, occur when two distinct genes affect the same phenotype in the same way.
One of the best examples of this would be the shape of the shepherd's purse pollen sac.
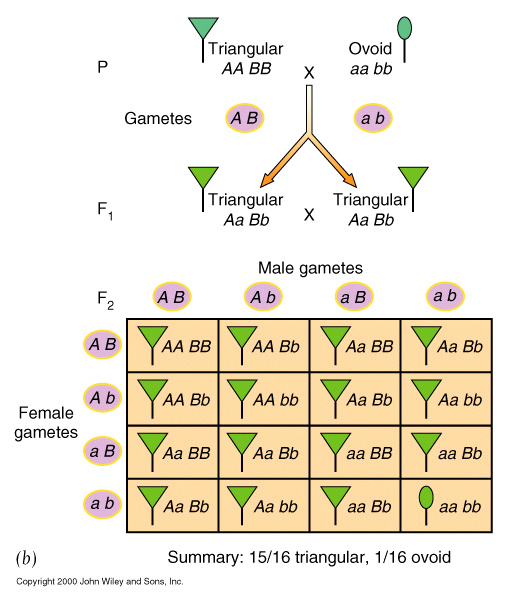
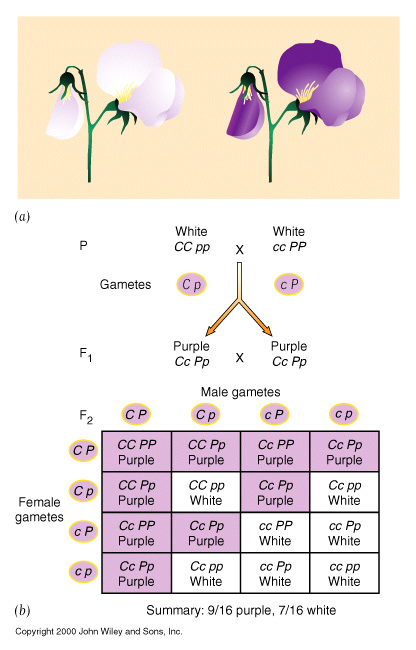
Complementary gene activity, occurs when two distinct genes affect the same phenotype in a complementary way, whereby the presence of BOTH dominant alleles are required to give a particular phenotype that is distinct from either being present by itself.
Polygenes also mediate quantitative inheritance.
Individual heritable characters
are often found to be controlled
by groups of several genes, called
polygenes.
Each
allele intensifies or diminishes the
phenotype.
Variation is continuous or quantitative
(“summation" or "adding
up” of all the traits).
Examples of continuous characters
are height, skin color, and (possibly)
intelligence.

| Incomplete dominance: | eg. Four o' clocks, carnations. BLENDING 1 : 2 : 1 |
| Codominance: | eg. M and N
blood groups on
chromosome 4 having
specific antigens
M and N
1 : 2 : 1 but where the heterozygotes (MN) gives rise to a distinct phenotype |
| Multiple alleles | ABO blood types, A and B are dominant to O , but A and B codominant to each other |
| Epistasis | essentially "eliminates"
or "masks" phenotypic
expression of other
genes,
eg. Labrador dogs fur colour, albinoism in mice |
| Lethal alleles | eg. Yellow colouration in mice fur. 2 : 1 . Pleiotropy(?) |
| Several genes / same character | Coat colour in mammals,
eg. mice A (agouti), B (black/brown), C, (colour) D (intensity), S. (distribution) genes |
| Complementary gene activity | eg. Pea plants, purple colouration. 9 : 7 |
| Duplicate gene activity | eg. Shepherd's
purse, Round over
narrow fruits, where
both A 1 - and A 2 - can cause heart shape 15 : 1 |
Nature
vs. Nurture?
When thinking about the importance
of either genotypes or phenotypes,
one must always consider the immense
importance of.....the environment , and how it can affect
the action(s) of any given gene.
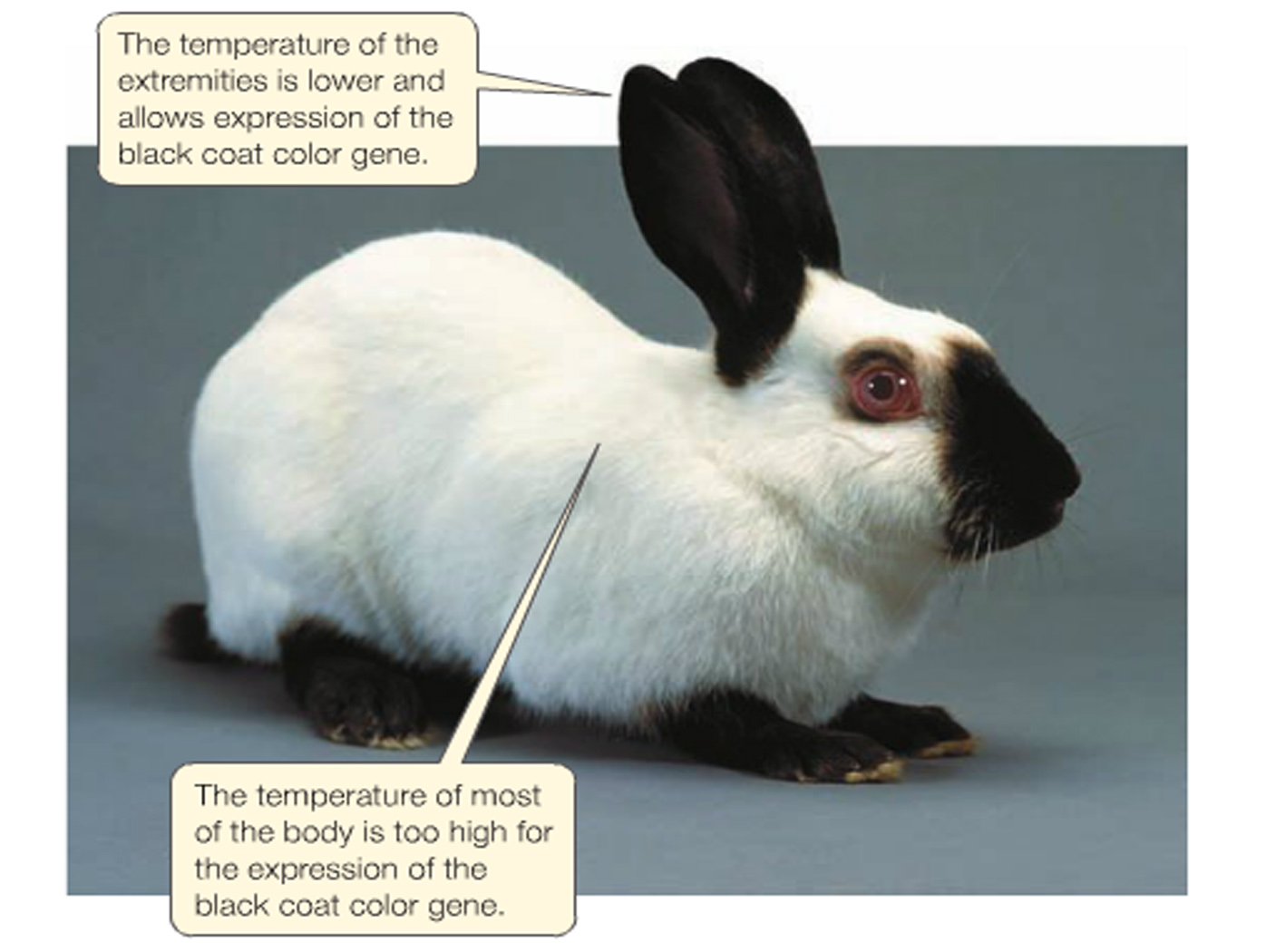
Variables such as light, temperature, and nutrition can dramatically affect the translation of a genotype into a phenotype.
For example, the darkness of the fur on extremities of a Siamese cat is affected by the temperature of that region. Darkened extremities normally have a lower temperature than the rest of the body.
Such colouration can be manipulated experimentally.
Gene linkage and the role of the sex chromosome:
Mendel did all his analyses with plants, which -like many other plants- exhibit BOTH male and female reproductive structures in the same adult plant (termed monoecious, or "all in one house").
{kind=link}
What about animals and plants that have individuals which are either one or the other sex (termed dioecious, or having “two houses”).
In most dioecious organisms, the sex of the organism is determined by differences in the presence/absence of a set of chromosomes, or the presence/absence of discrete chromosomes.
In honeybees, eggs are either fertilized -and become diploid females- or they are not fertilized -and become haploid males, drones.
In Mammals, such as humans...
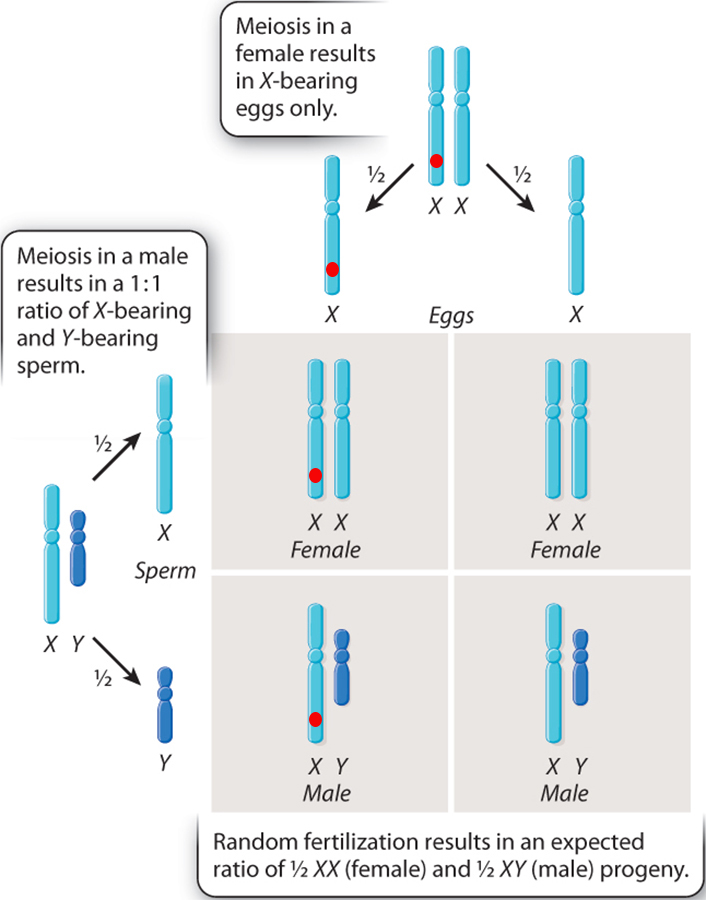
Humans have different sex chromosomes, X and Y. Males have X and Y; females have X and X.
Again, the sex of the offspring is also determined by the sperm.
If a sperm with an X chromosome reaches the egg, the resulting offspring will be female (XX). If a sperm with a Y chromosome reaches the egg, the resulting offspring will be male (XY).
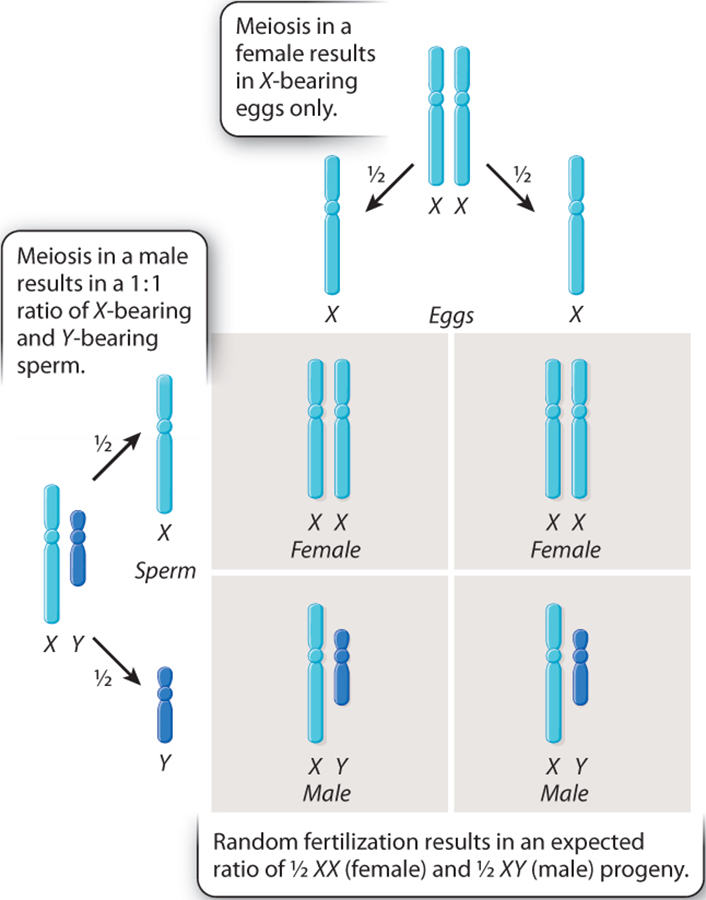
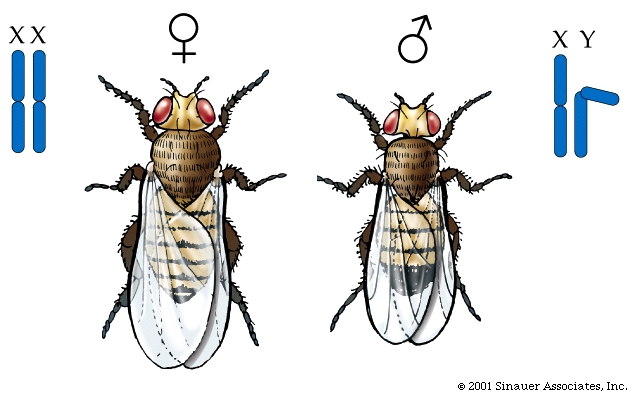
In the Fruit fly (or Drosophila melanogaster) chromosomes also have distinct X and Y chromosomes, wherein the male is XY and the female is XX. As a consequence, like humans, the male fruit flies are said to have a set of paired "autosomal" chromosomes and then one X and one Y chromosomal. They are, therefore said to be hemizigous.
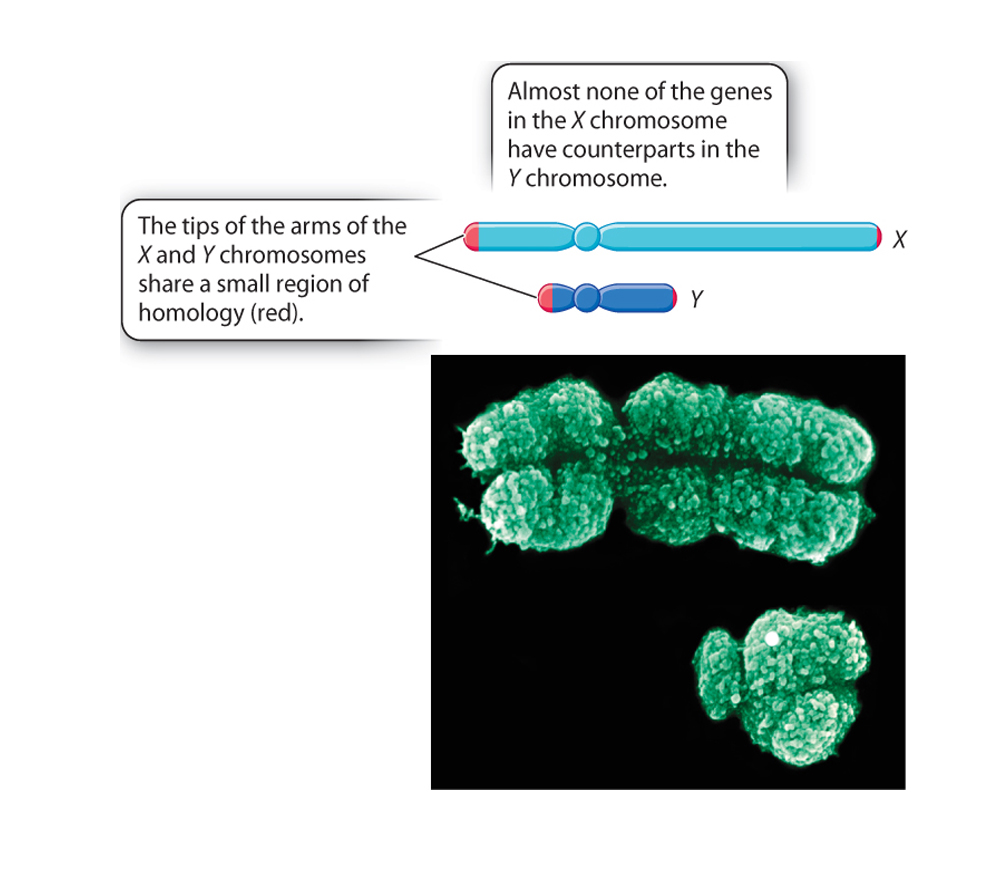
In mammals the X and Y chromosomes have different functions
The gene that determines maleness in mammals was identified by studying people and animals with chromosomal ploidy abherations... as, unlike most autosomal ploidy variations, mammals can handle some varaiation in the numbers of X and Y chromosomes.XY females often have a piece of the Y chromosome missing, whereas the XX males have a piece of a Y attached to the X.
The fragment missing from the Y chromosome in XY females or that needs to be present on the X chromosome in XX males contains the maleness-determining gene.
The gene was named SRY (for Sex-determining Region on the Y chromosome).
The SRY gene codes for a functional protein (TDF or Testis Determining Factor) involved in primary sex determination.
A gene on the X chromosome called DAX1 produces an "anti-testis" factor. The SRY gene product in a male inhibits the gene DAX1, and consequently no "male-specific" inhibitor is made.
Secondary sexual traits like breast development, body hair, and voice are also influenced by hormonal levels of key sex hormones -such as testosterone and oestrogen.
The presence of Sex chromosomes allows for a special type of genetic inheritance to be analyzed..... sex-linked inheritance. In essence, while a female can be heterozygous for a particular gene that is present on the X chromosome, her male offspring will be hemizygous for that particular trait.
if the allele on the X chromosome is recessive, whil it may be "masked" in a heterozgous female it will always show through in her male offspring.
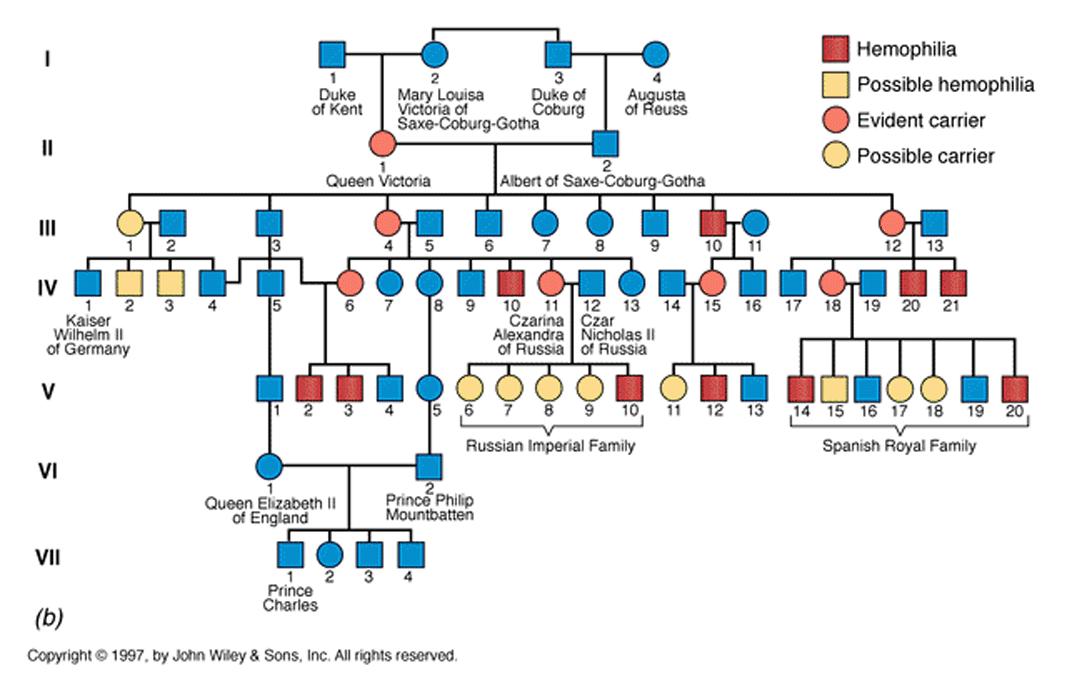
Consequently it is relatively easy to trace an X-linked trait that has an overt phenotype. One such trait, which has become somewhat infamous among genticists (especially in England because there are lots of data available with respect to lineage and heritable traits) is the passage of Haemophaelia within the English royal family.
On a less morbid note about another type of X-linked trait Red, Green Colour blindness.... true story!



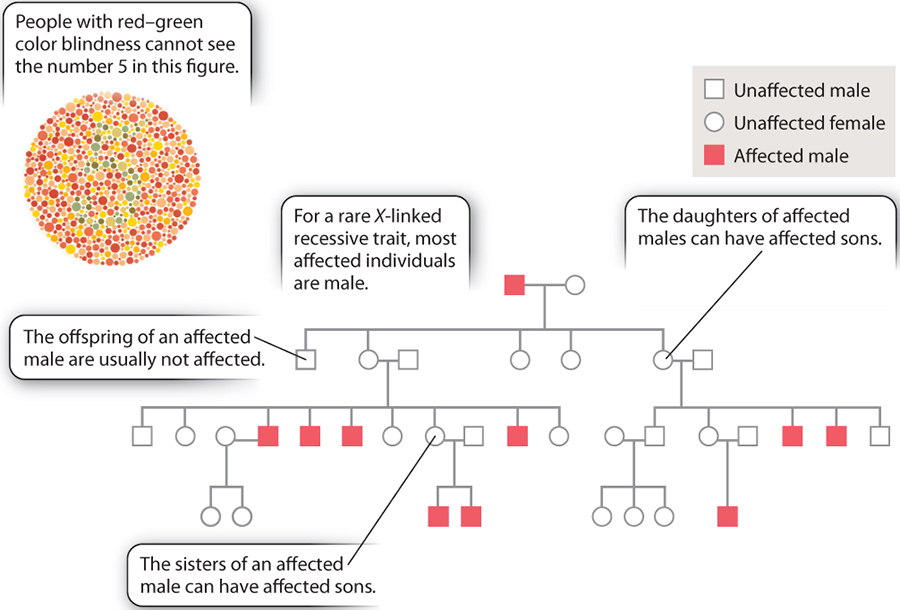

While the X and Y chromosomes normally behave according to the laws of meiotic cell division (and thus according to Mendelian genetics) the ability to "visualize" the presence of each of the alleles that is located on either of the X-chromosomes of a mother (by observing the different phenotypes) challenge the ubiquity of Mendel's 2nd law, or in other words...
Mendel's 2nd Law DOES NOT ALWAYS APPLY
Remember, that ......to equate Mendel with meiosis we had to invoke the role of "chiasmata" an their occurance between two gene loci on adjacent chromatids in paired, "bivalent" chromosomes during prophase I of meiosis.
We now knowthat this CANNOT occur for X and Y chromosome... OK, but what about ALL the genes on the other autosomal chromosomes. Again, I glibbly stated that, if the probability of a single chiasma forming between two genes is "one" (i.e. a certainty), then the assortment of alleles would, in essence, be the same as if they were on separate chromosomes (i.e. random).
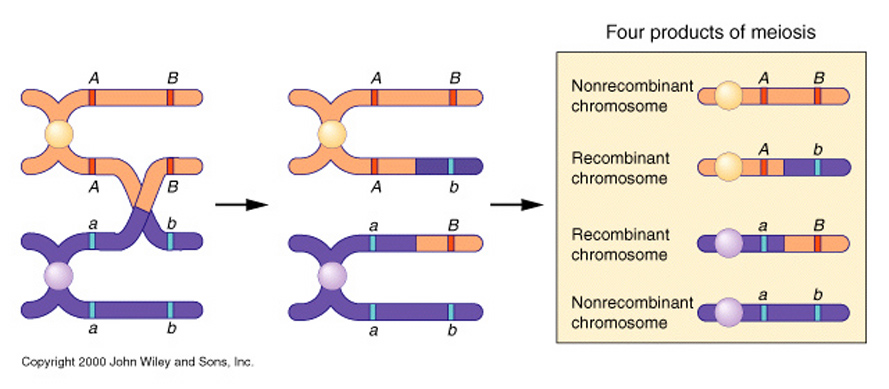
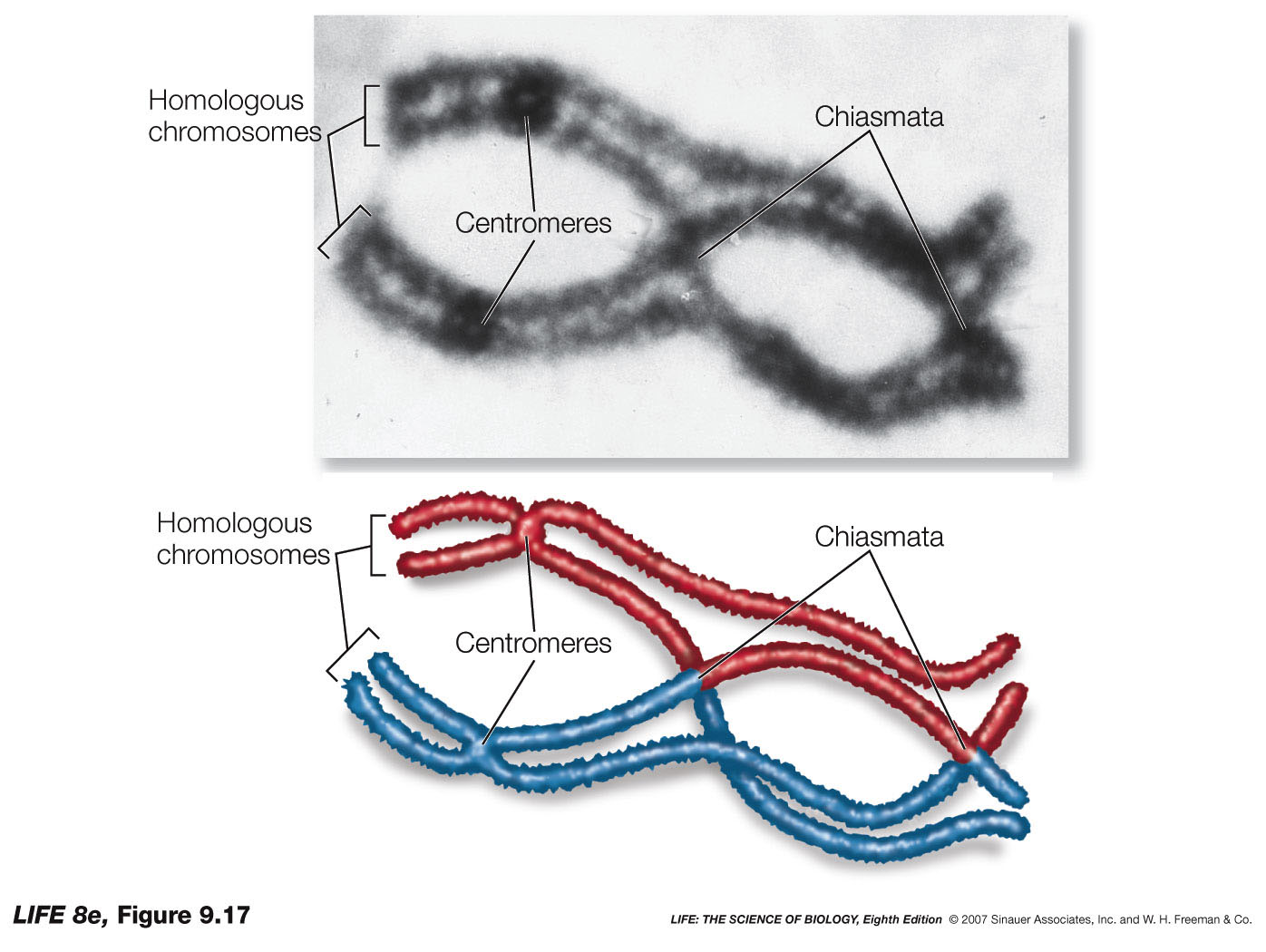
But, what happens if there are no chiasmata formations?
Then the genes on the same chromosome cannot assort randomly, which would be in marked contrast to Mendelian expectations.
However, this phenomenon can and does happen.
Such aberrations were first hypothesized to exist by Bateson and Punnett, who observed some curious "asssociations" of heritable traits. However, it was really verified by an American geneticist, Morgan (1909) who was working on chromosomally-linked genes that resided on the X chromosome of the fruit fly Drosophila melanogaster.
So, what happens if there are NO CHIASMATA formations between homologsous chromosomes once they come together in .... phase of Meiosis?
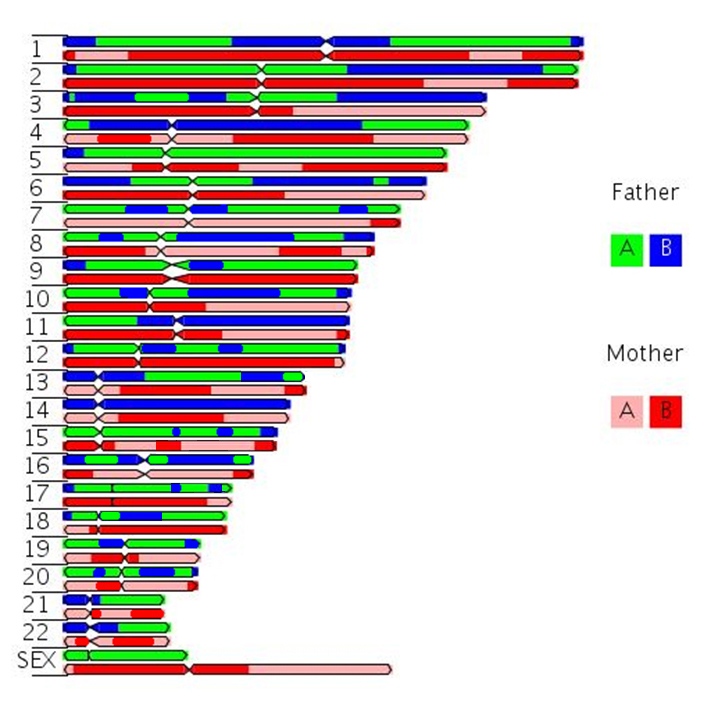
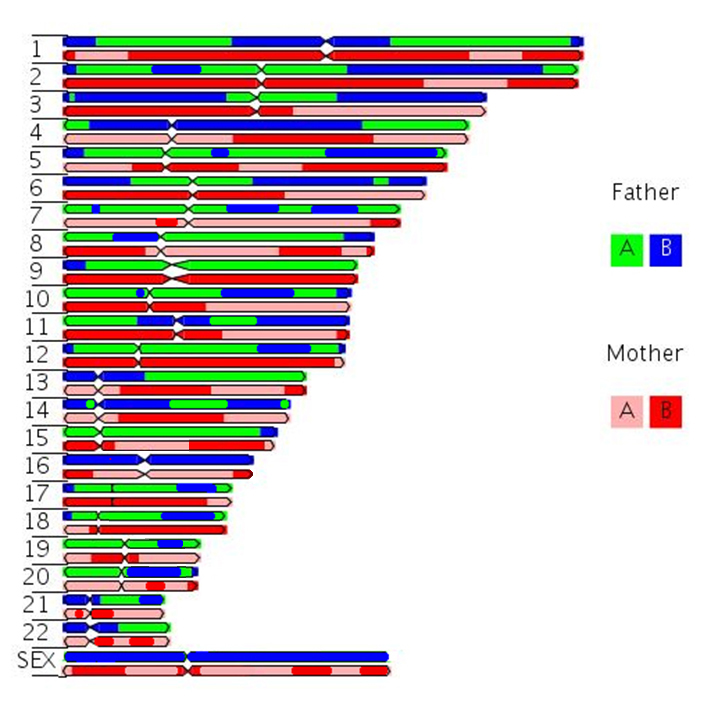
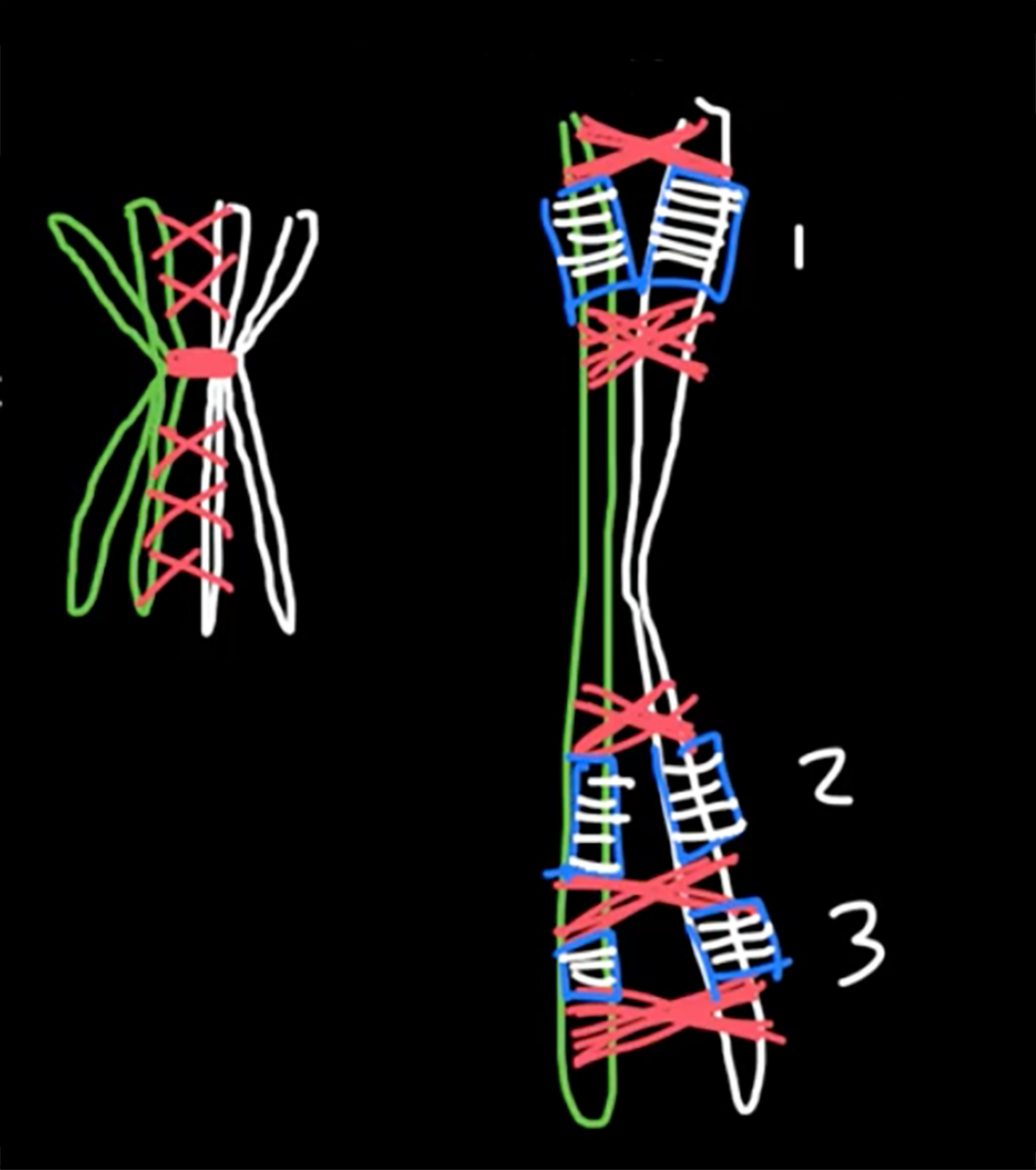
Morgan explained this apparant anomally by proposing that the two loci were present and "linked" on the same chromosome, and that any variation from the parental "linkage" must have occurred through the exchange of genetic material by some type of "crossover events" occurring between these two loci.....
While not common, this "linkage" phenomenon can also happen -even with some genes on the autosomal chromosomes.
Again, looking back to previous lectures, I glibbly stated that, if the probability of a chiasma forming between two genes is "one" (i.e. a certainty), then the assortment of gene pairs on the same chromosome would (in essence) be the same as if they were on separate chromosomes (i.e. randomly assorted -as predicted by Mendel).
Such aberrations were first hypothesized to exist by Bateson and Punnett, who observed some curious "asssociations" of heritable traits. However, it was really verified by an American geneticist, Morgan (1909) who was working on chromosomally-linked genes that resided on the X chromosome of the fruit fly Drosophila melanogaster.
To me it is a curious irony that the vindication of Mendel/Sutton-Boveri's ideas that merged Mendelian genetics with cytological analysis of chromosomes, came about through research on sex-linked genes (that don't obey mendel's 2nd law) because (remember) Mendel's early experiments, were carefully chosen to be sex independent, i.e. he only worked with autosomal genetic traits in a monoecious pea plant
