- Home
Welcome !
Welcome to john houghton's home page for his biology courses. This site is designed as a hub for curating and sharing lectures, course syllabi, assignments, and links to relevant resources. Use the menu bar at the top of the screen to navigate through the site.
(Please note: this page is currently under construction.)

- BIOL 2107
Fall '22 CRN87989
Lectures: (1)
- Courses

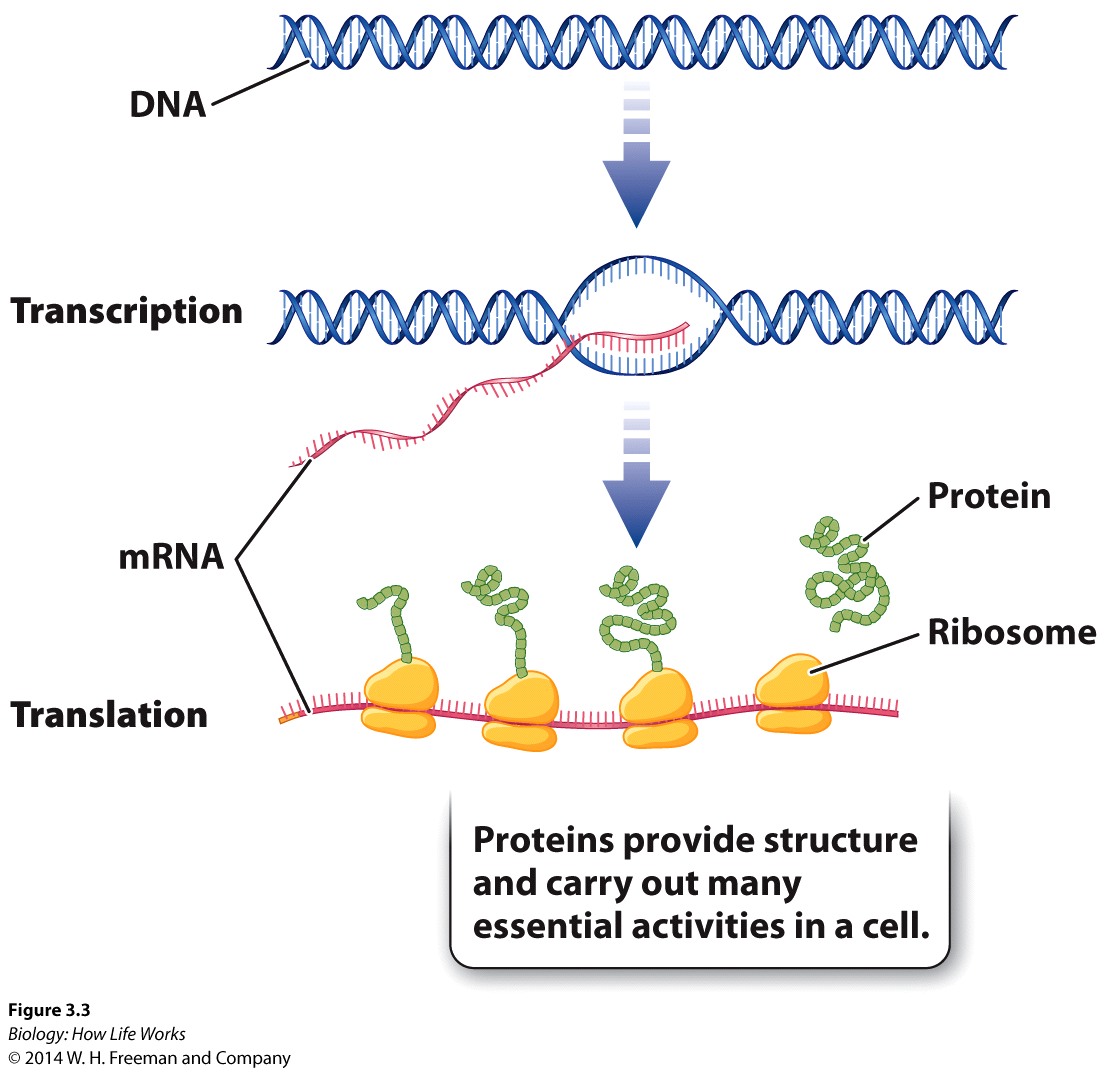
Lecture 22: Central Dogma: A little bit of History: ![]()
![]()
DNA:base-pairing -Animation DNA -Animation
The CENTRAL DOGMA.....
DNA: The Genetic Material. Well we all know that, but (curiously) for the longest time we didn't. During the first half of the twentieth century, the structure that carried hereditary information from generation to generation was generally assumed to be a protein.
The diversity and specificity of proteins seemed appropriate for genetic material.
DNA seemed to be just too simple to carry complex information.
Circumstantial evidence, however, began to point to DNA as being the true genetic material.
DNA was found in the nucleus, which was already known to carry genes.
A dye that bound to DNA showed that the amount of DNA in somatic cells was twice that in eggs or sperm, as would be expected from Mendel's discoveries. But all this DNA was wrapped up in a protein:DNA complex called chromatin, which comprised the chromosome. Indeed, every other "-some" known to man is always a complex of nucleic acid and protein. So, the question obviously became obvious, which of this -somal complex provided the structure, and which provided the "true" genetic material.
Previously, in 1928, the English physician Frederick Griffith had made a landmark discovery about heredity while looking for a vaccine against Streptococcus pneumoniae one of the bacteria that causes pneumonia in humans.
Griffith worked with two different strains of the bacterium.
The S strain produced shiny, smooth colonies when grown in the laboratory.
The R strain produced coloniesthat looked rough.
The S strain was virulent (mice injected with the S strain died within a day); a capsule around the S strain bacteria protected them from the host's immune system.
The R strain lacked this "capsule" and was avirulent.
Griffith heated some S strain bacteria to kill them, then injected the bacteria into mice.
The heat-killed bacteria did not kill the mice......... indicating that the bacteria had to be ALIVE to cause death to the host.
However, a mixture of heat-killed S strain bacteria and living R strain bacteria did kill the mice. Huh??
Moreover, Griffith found that all the live bacteria in the hearts of the mice that had been killed in this way were now S strain.
Conclusions: that some of the living R strain bacteria had been genetically transformed/modified by the presence of the heat-killed S strain bacteria. But what?
Further tests demonstrated that some substance from the dead S strain bacteria could cause a heritable change in the R strain bacteria, thus defining the 'transforming principle'suggesting that what ever was transferred to the R Strain must have carried heritable information and thus was the genetic material.
Oswald T. Avery and colleagues spent several years identifying the transforming principle by a process of elimination.......DNA
This work was published in 1944, but was not immediately appreciated.
Viral/phage replication experiments confirmed that DNA is the genetic material
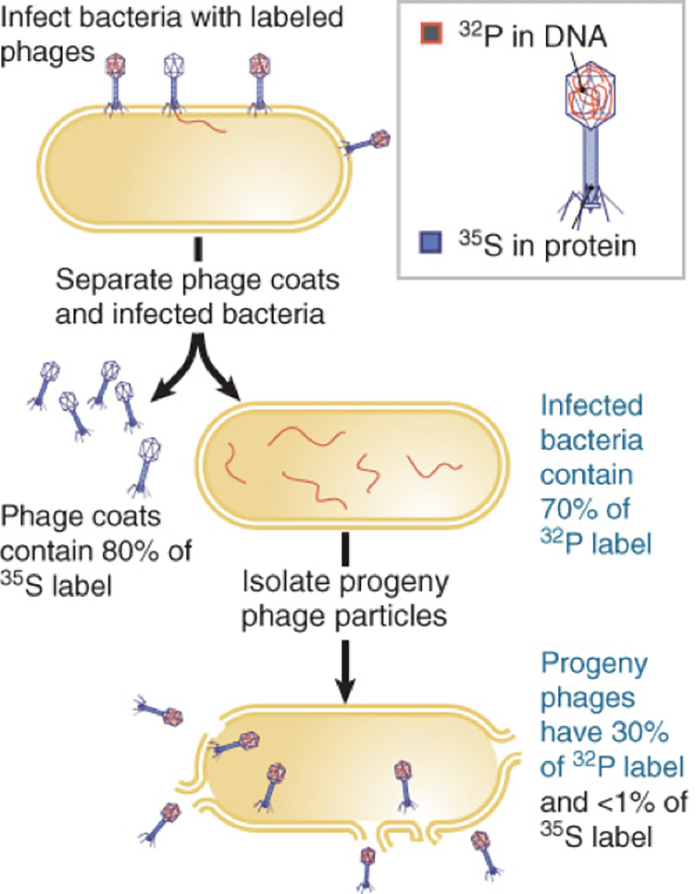
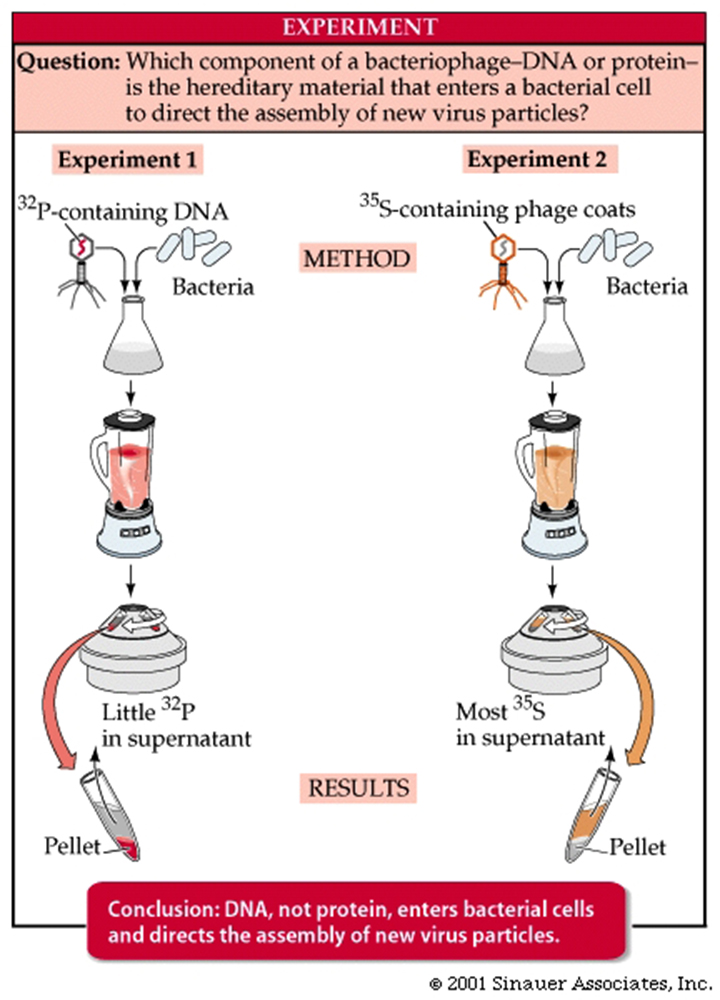
In 1952, Alfred D. Hershey and Martha Chase performed experiments confirming that DNA is the genetic material.
The T2 bacteriophage, a bacterial virus that attacks the bacterium E. coli, consists almost entirely of a DNA core packaged within a protein coat.
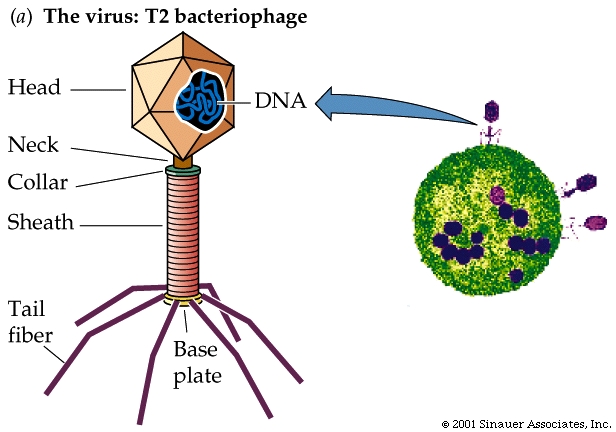
When a T2 bacteriophage attacks a bacterium, part but not all of the virus enters the bacterial cell.
The Hershey-Chase experiment determinedwhich part of the virus entered the bacterium.
They labelled some phage with radioactive sulphur. (remember sulphur can be present in proteins BUT NOT in DNA).
Other bacteriophage they labelled with radioactive phosphorus. Phosphorus is present in DNA but absent from most proteins.
So, using these two radioactive mechanisms to discriminate between the genetic source, viruses with labeled sulphur or phosphorus were allowed to infect non-radioactive E. coli cells.
By quickly putting the infected bacteria into a blender (to remove any phage that may have been attached to the outer membranes) and then separating the cells from the phage by centrifugation, Hershey and Chase were able to demonstrate that the labeled sulphur (and thus the viral protein) had separated from the bacteria, but that the labelled phosphorus (and thus the viral DNA) remained with the bacteria.
Experiments on later generations of bacteria confirmed that the labeled phosphorus remained with subsequent generations while any residual labelled sulphur was quickly lost.
The Structure of DNA
Scientists now set out to determine the structure of DNA, hoping to find the answers to two questions:
How is DNA replicated between nuclear divisions?
How does DNA cause the synthesis of specific proteins?
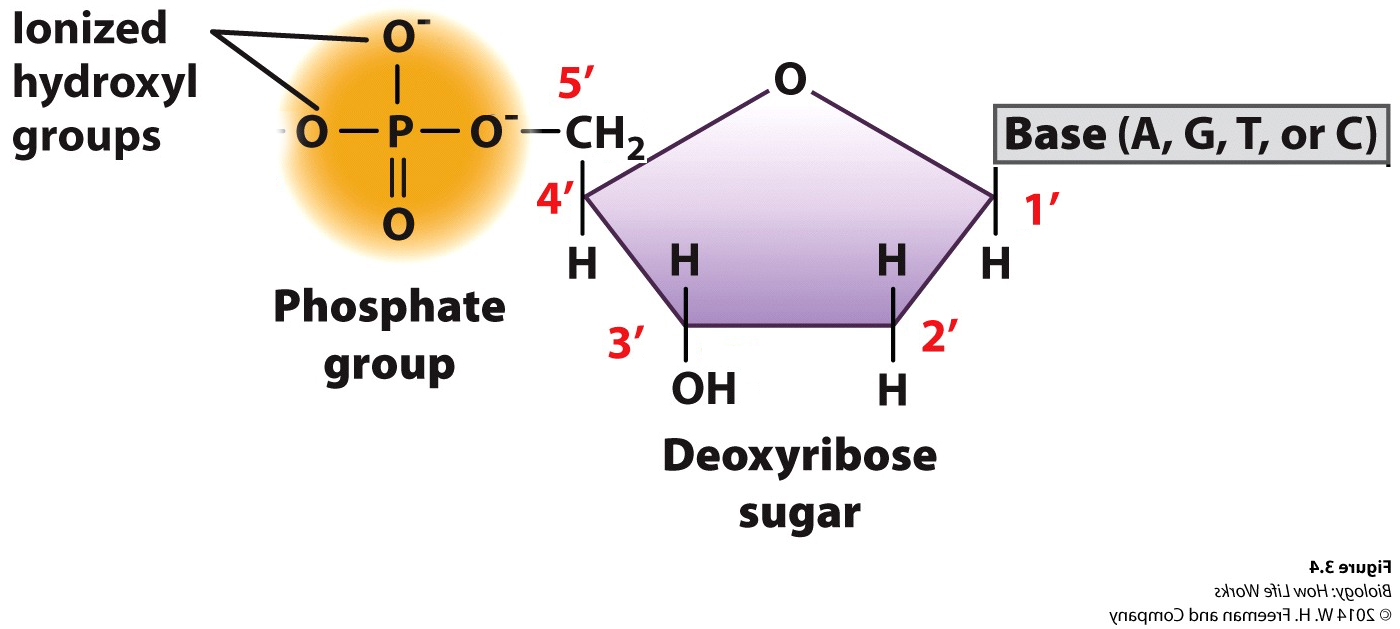
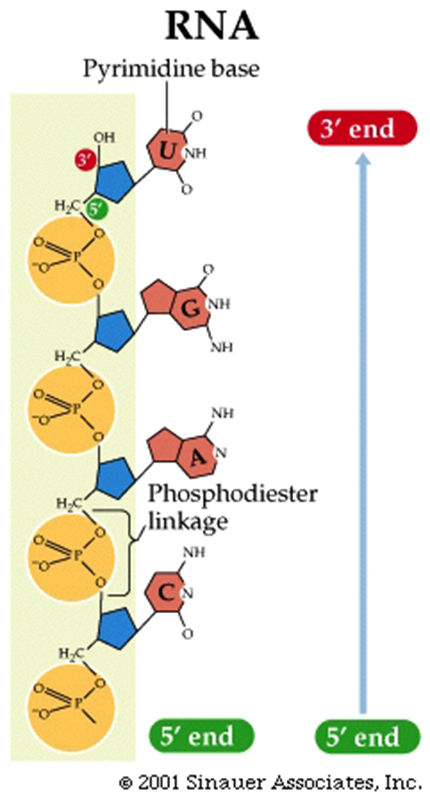
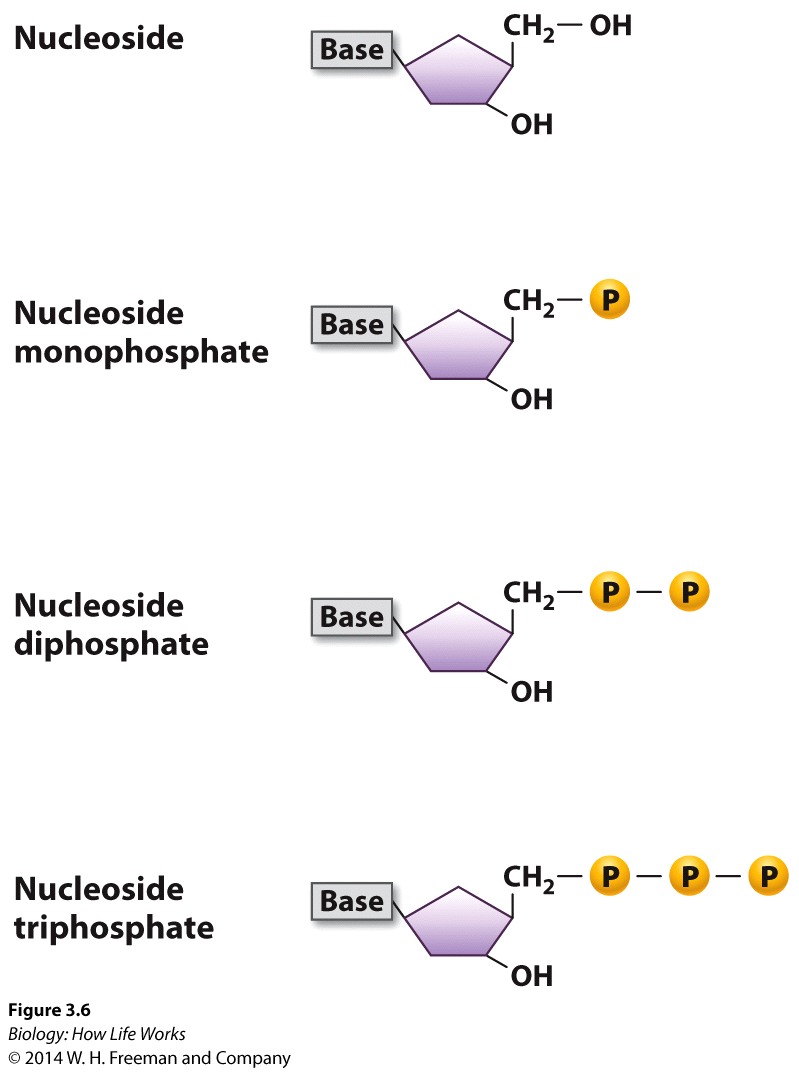
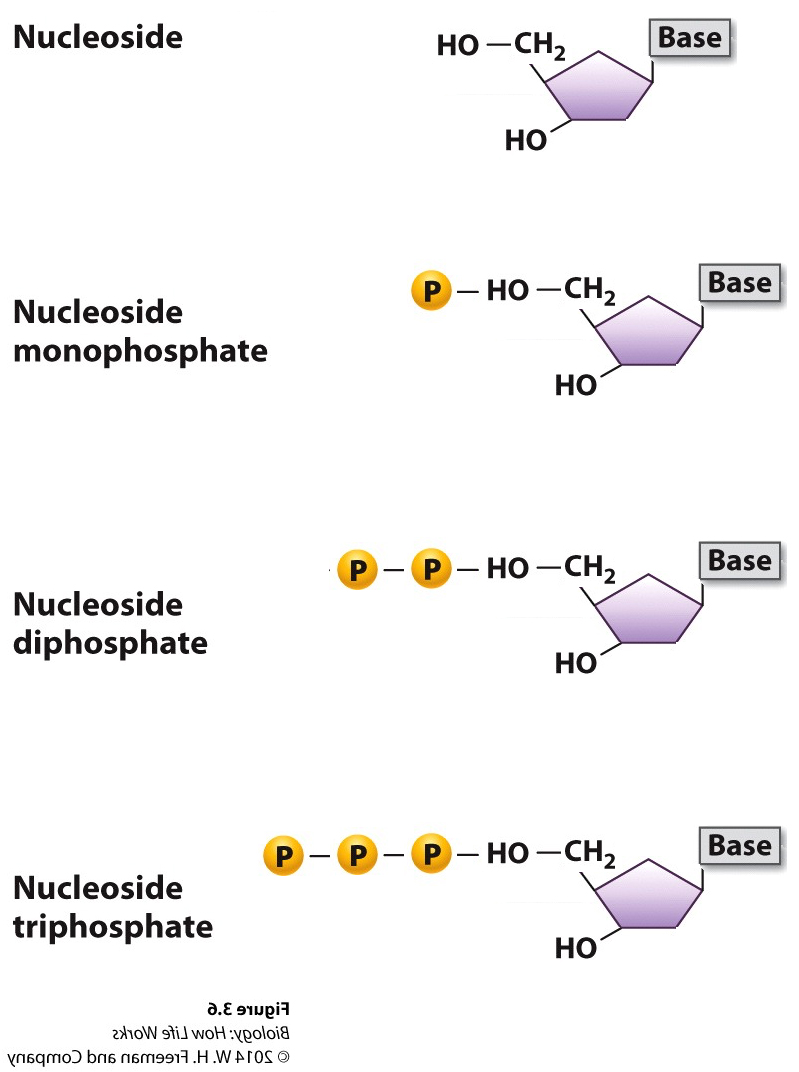
DNA was (and still is) composed of: Bases; Adenine, Thymine, Guanine and Cytosine, plus PO42- and a deoxy-ribose sugar moiety. As to how these nucleotides were arranged in space their spatial interactions….who knew?
The four nucleotides that make up DNA differed only in their nitrogenous bases.
The two purines (adenine and guanine) and two pyrimidines (cytosine and thymine).
In 1950, Erwin Chargaff noted that in DNA from all species tested, the amount of purines (adenine and guanine) always equaled the amount of pyrimidines (cytosine and thymine. Not only this, but, curiously, the amount of adenine (in purified DNA) always equaled the amount of thymine, and the amount of guanine equaled the amount of cytosine.
X-ray crystallography provided some of theoretically important clues to DNA structure.
The positions of atoms in a crystalline substance can be inferred from the pattern of diffraction of X-rays which passed through it.
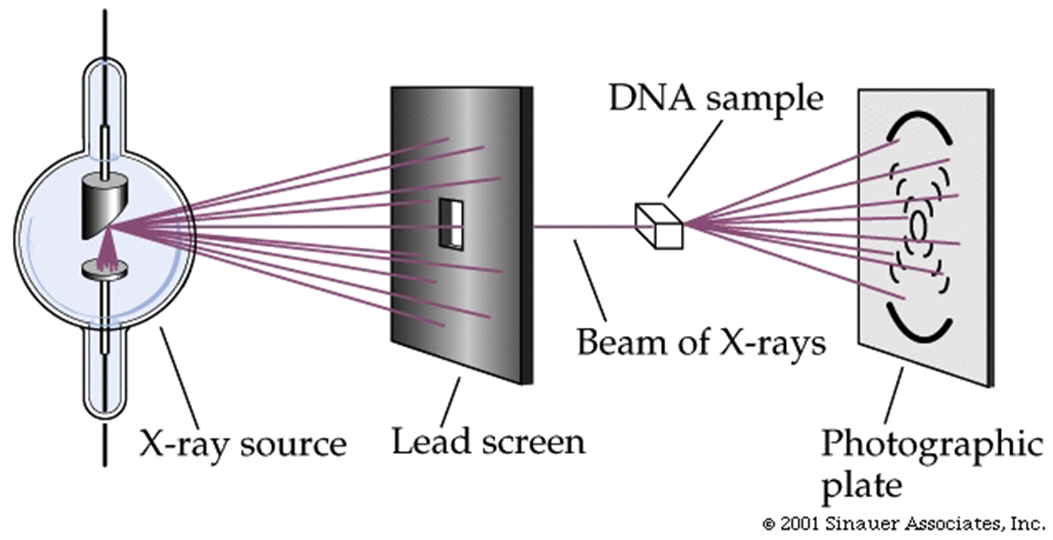
In the early 1950s, many skilled X-ray crystallographers tried but failed to glean information from X-ray diffraction patterns of DNA.
Rosalind Franklin, building on the previous work of Maurice Wilkins (her mentor), finally was able to provide key information about the structure of DNA based on its x-ray crystallography diffraction pattern.
From this (stolen?) information Watson and Crick imagined and then described the double helix, using the technique of model building to test their hypotheses.
The results of X-ray crystallography convinced them that the DNA molecule was helical.
X-ray crystallography also provided the values of certain distances between key elements of the helix.
Density measurements and earlier models pointed to a structure with two polynucleotide chains running anti-parallel to each other.
Although there have been modifications, the principle features of the model they built in 1953 have remained essentially unchanged.
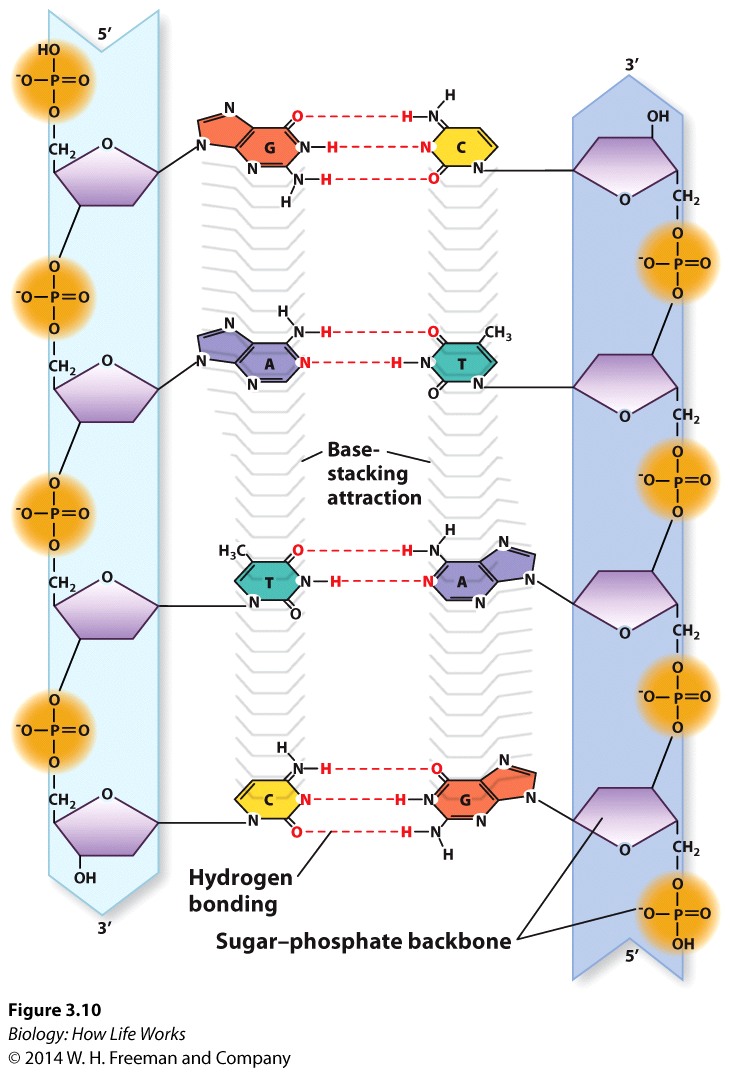
The DNA molecule is a double-stranded helix.The diameter of the DNA molecule is uniform.
The twist in DNA is "right-handed" (the twist is in the same direction as the thread on most screws).
The DNA molecule is a double-stranded helix.The diameter of the DNA molecule is uniform, and easily interpreted by proteins that "patrol" the DNA to maintain it's integrity.
The twist in DNA is right-handed (-the same direction as the threads on most screws)...right handed double-helix, although the "threading" is not always the same.....B-form HelixThe two strands run in different directions (they are antiparallel ).
The sugar-phosphate backbones of each strand coil around the outside of the helix.
The nitrogenous bases all point toward the center of the helix.
Hydrogen bonds -a type of attractive intermolecular force that exists between two partial electric charges of opposite polarity- between complementary bases hold the two strands together.
A always pairs with T (two hydrogen bonds).
Consequently, each base pair has one purine and one pyrimidine, so the diameter of the double helix remains constant.
As the genetic material of the cell, DNA must perform four important functions:
It must be able to store all of an organism's genetic information.It must be susceptible to mutation.It must be precisely replicated in the cell division cycle.It must be expressible -giving rise to the phenotype.As we have seen, the simple, double-helical structure of DNA, with the two strands linked by complementary base pairs, readily lent itself well to the first two requirements of its function.
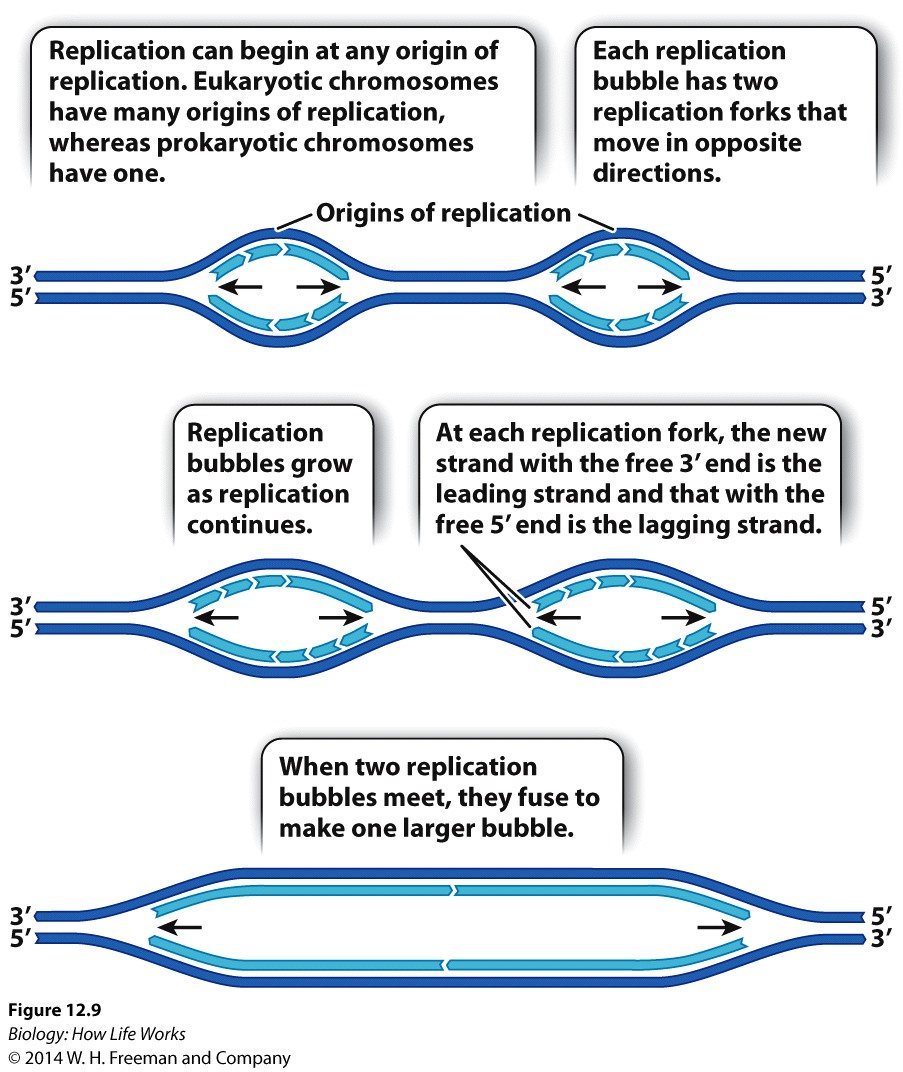
Replication of DNA is semiconservative, with both strands of DNA acting as templates for each "growing strand of DNA". But replication of DNA is also directional (always elongating in the 5' -> 3' direction.
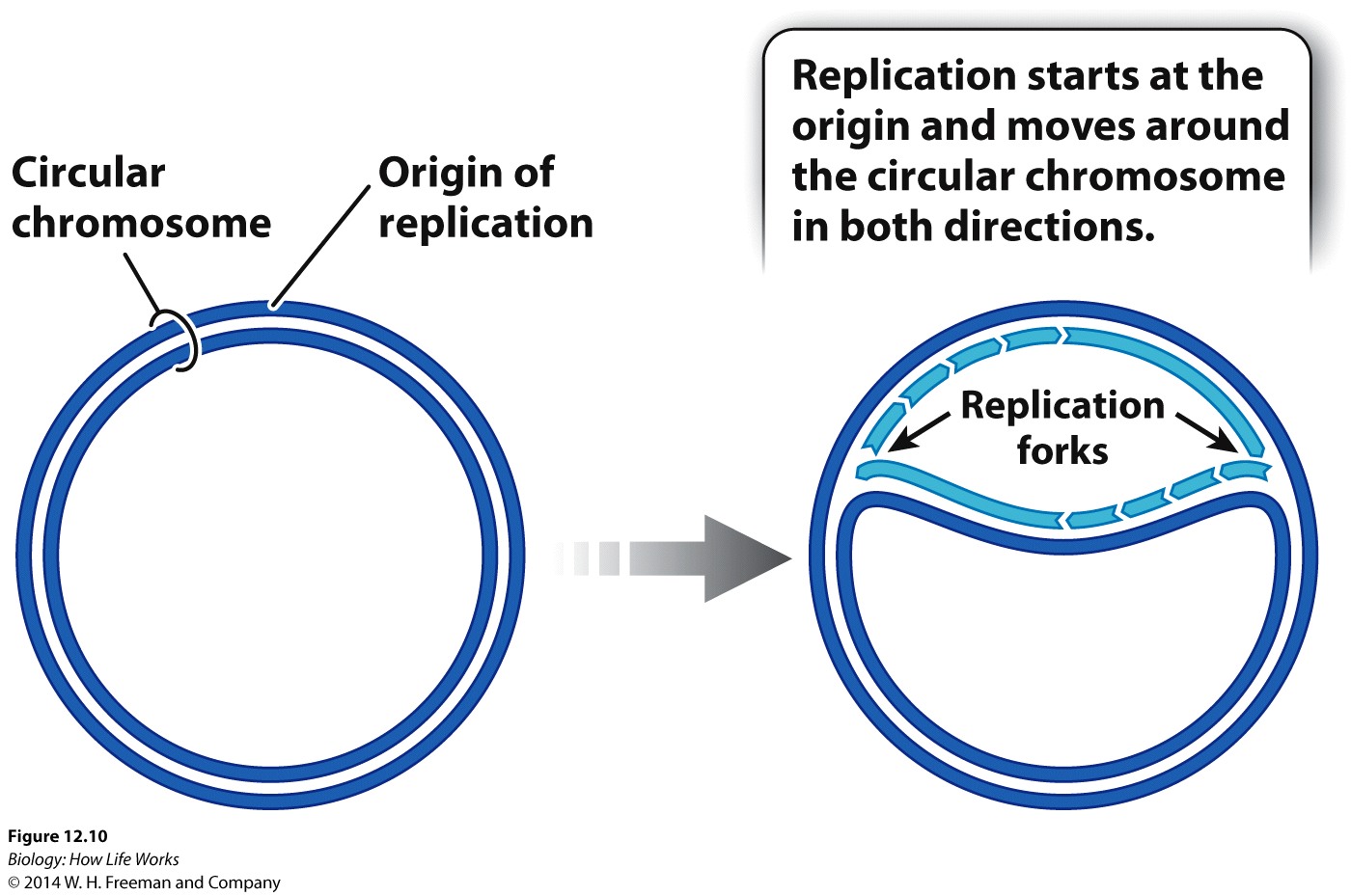
In eukaryotes, large chromosomes can have hundreds of origins of replication. Consequently, in eukaryotes, DNA replication occurs at many different sites simultaneously.
In both eukaryotes and prokaryotes, DNA is replicated by a large enzyme complex, called DNA polymerase or DNA Replicase, as it catalyzes the reactions of DNA replication.
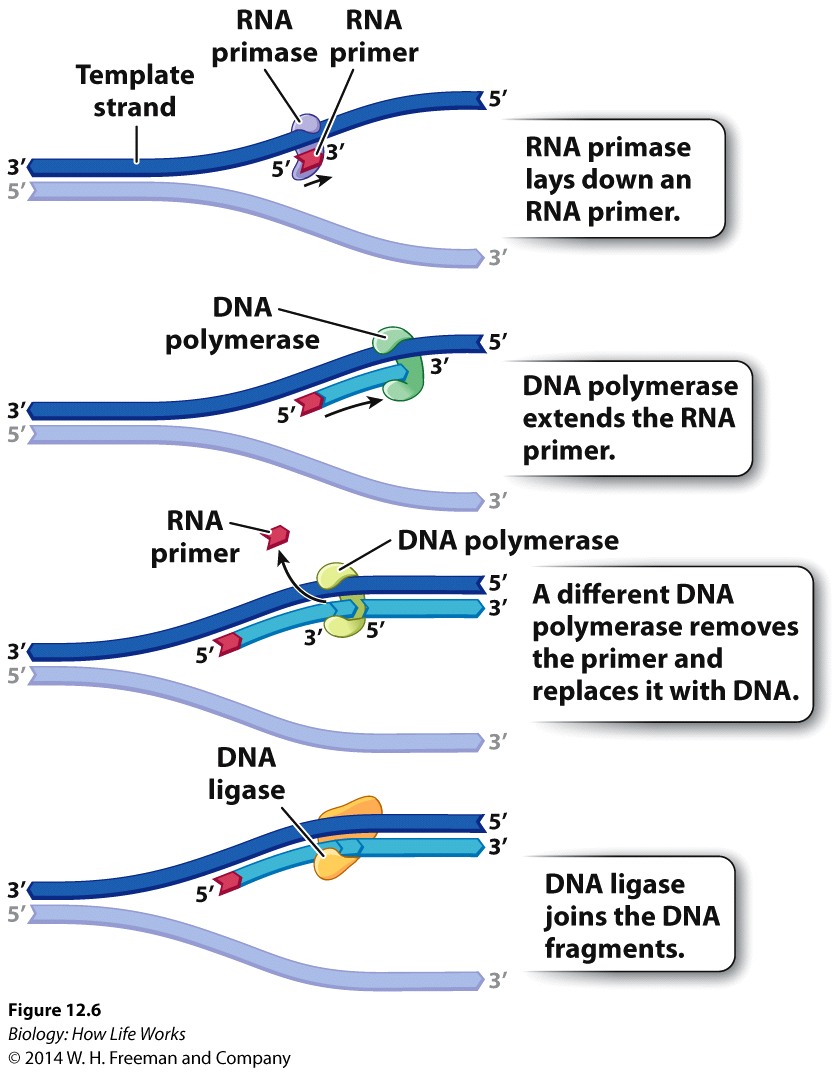
Because DNA polymerases are so faithful, the process of DNA replication has to begin from an existing 3'OH group.... As a result Replication of chromosomes by the Major replicase (DNA Polymerase) ALWAYS needs a "primer", which is a short strand of RNA complementary to the DNA template strand.
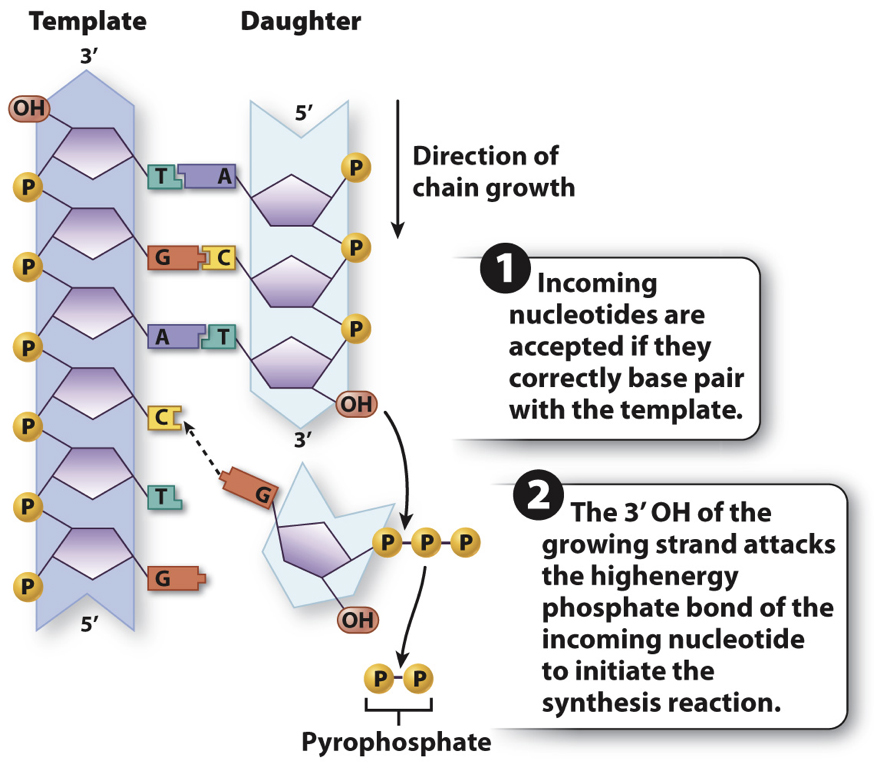
An enzyme called a primase makes the RNA primer strand.
The RNA primer is later degraded and replaced with DNA, so the final DNA molecule has no RNA.But we now have a problem......!!!!
So both strands cannot be synthesized in exactly the same way.
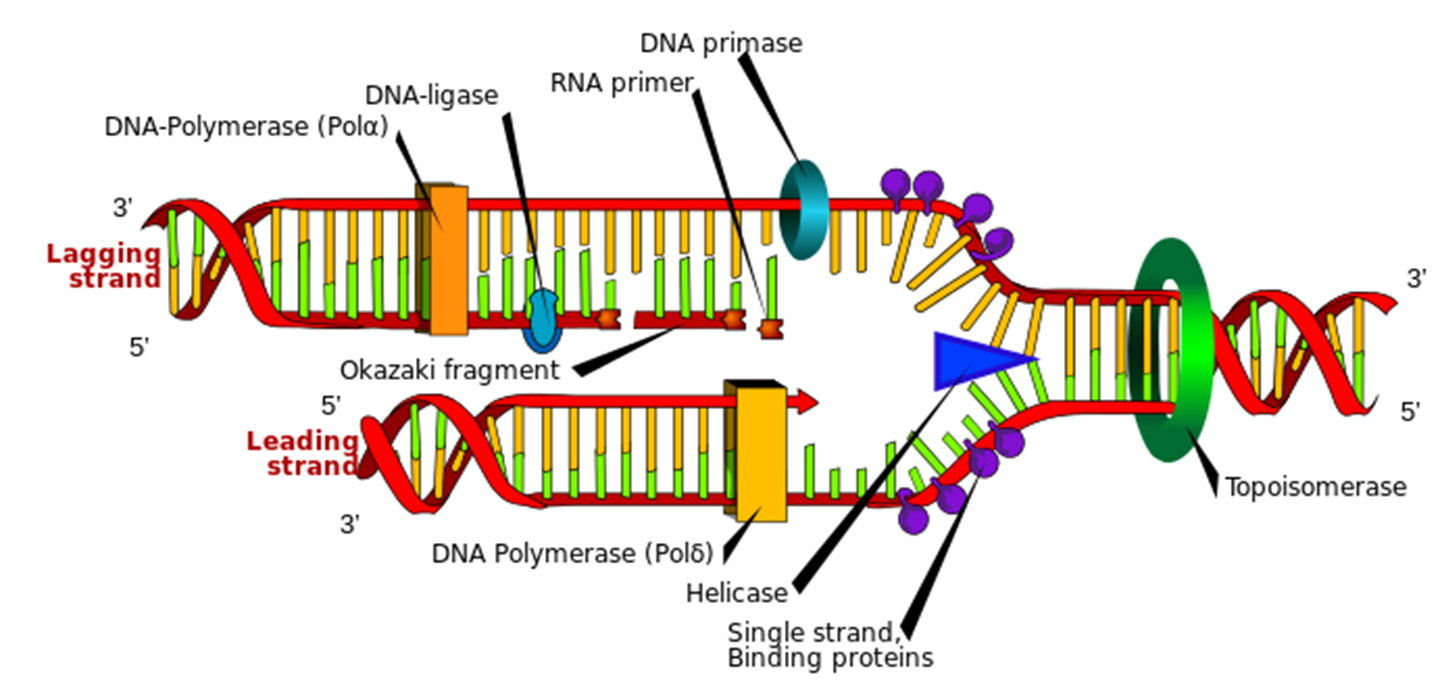
DNA replication involves the activities of several "key" proteins,each with different roles:
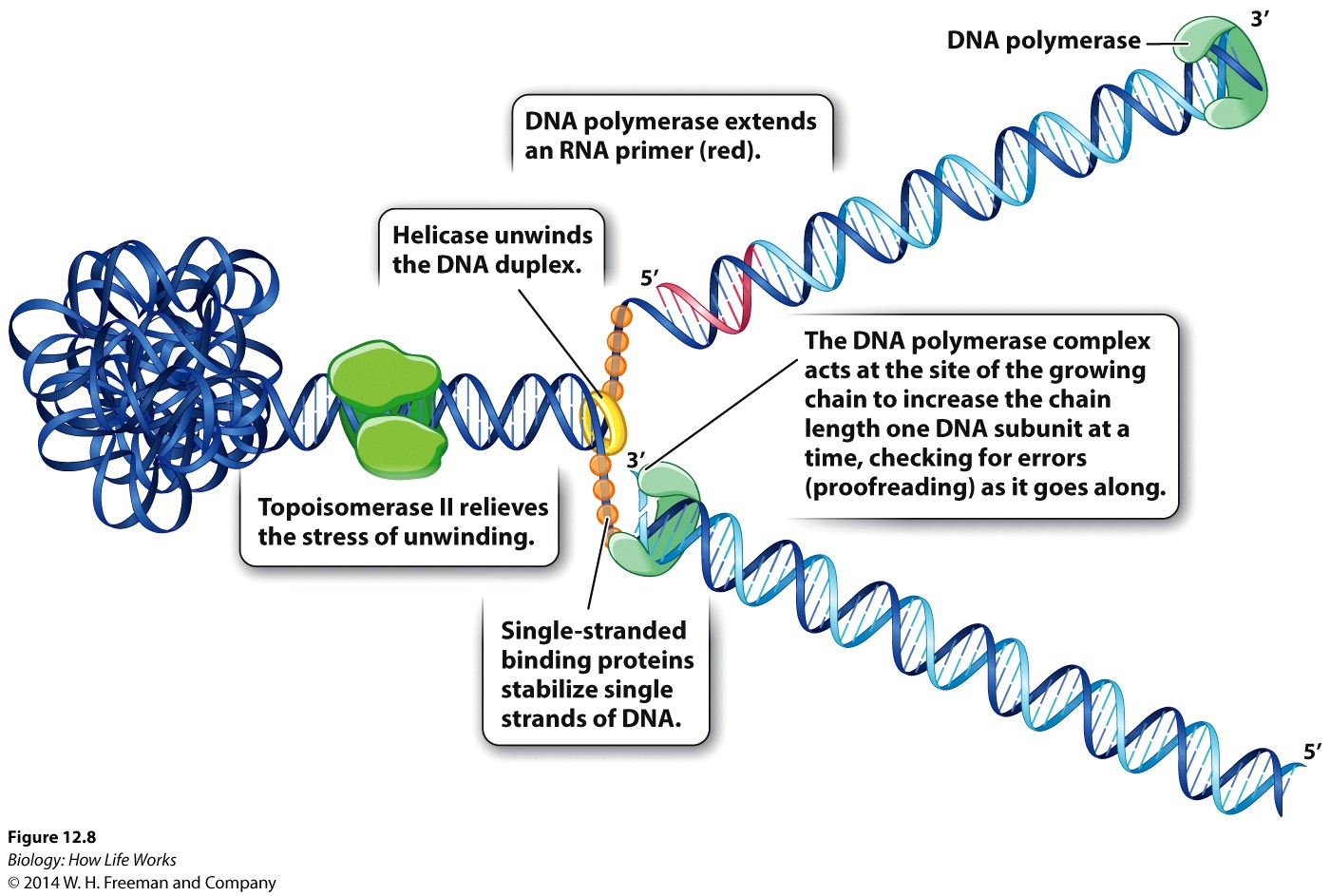
DNA helicase & topoisomerase: which denature and untangle the double helix.
Single-strand binding proteins: keeps the two strands separate.
RNA primase: makes a primer strand that serves as a starting point for replication.
DNA polymerase: itself adds complementary nucleotides to the growing strand, proofreads the DNA, and repairs it.
DNA ligase: seals up breaks in the sugar–phosphate backbone.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}