

- Home
Welcome !
Welcome to john houghton's home page for his biology courses. This site is designed as a hub for curating and sharing lectures, course syllabi, assignments, and links to relevant resources. Use the menu bar at the top of the screen to navigate through the site.
(Please note: this page is currently under construction.)

- BIOL 2107
Fall '23 CRN86772
Lectures: (1)
- Courses
BIOL 2107 Principles of Biology I

| Lecture 11: Mendelian Genetics I |
Review Cell Division....
{kind=link}

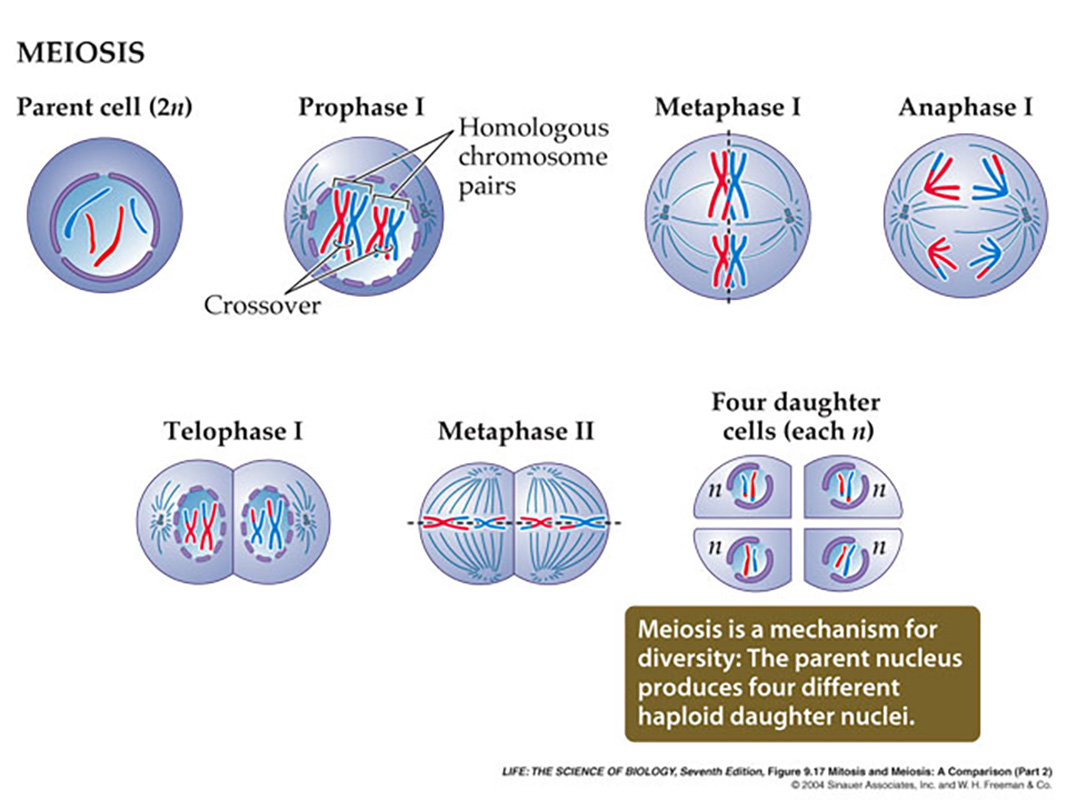
While meiosis provides the required genetic constancy from generation to generation, as I mentioned, it also provides for a considerable degree of variation in the offspring -without introducing mutations or errors in the distribution of the chromosomes within any of the gametes.
For the sake of completion, as to how and when the male and female gametes develop is both species and sex dependent.
Sometimes, however, the incredible choreography -that is cell division- doesn't always go according to plan.
Nondisjunction is the most commonly occuring error, in ths regard, wherein either homologous chromosomes fail to separate during anaphase I, or sister chromatids fail to separate during anaphase II......resulting in a condition known as aneuploidy.
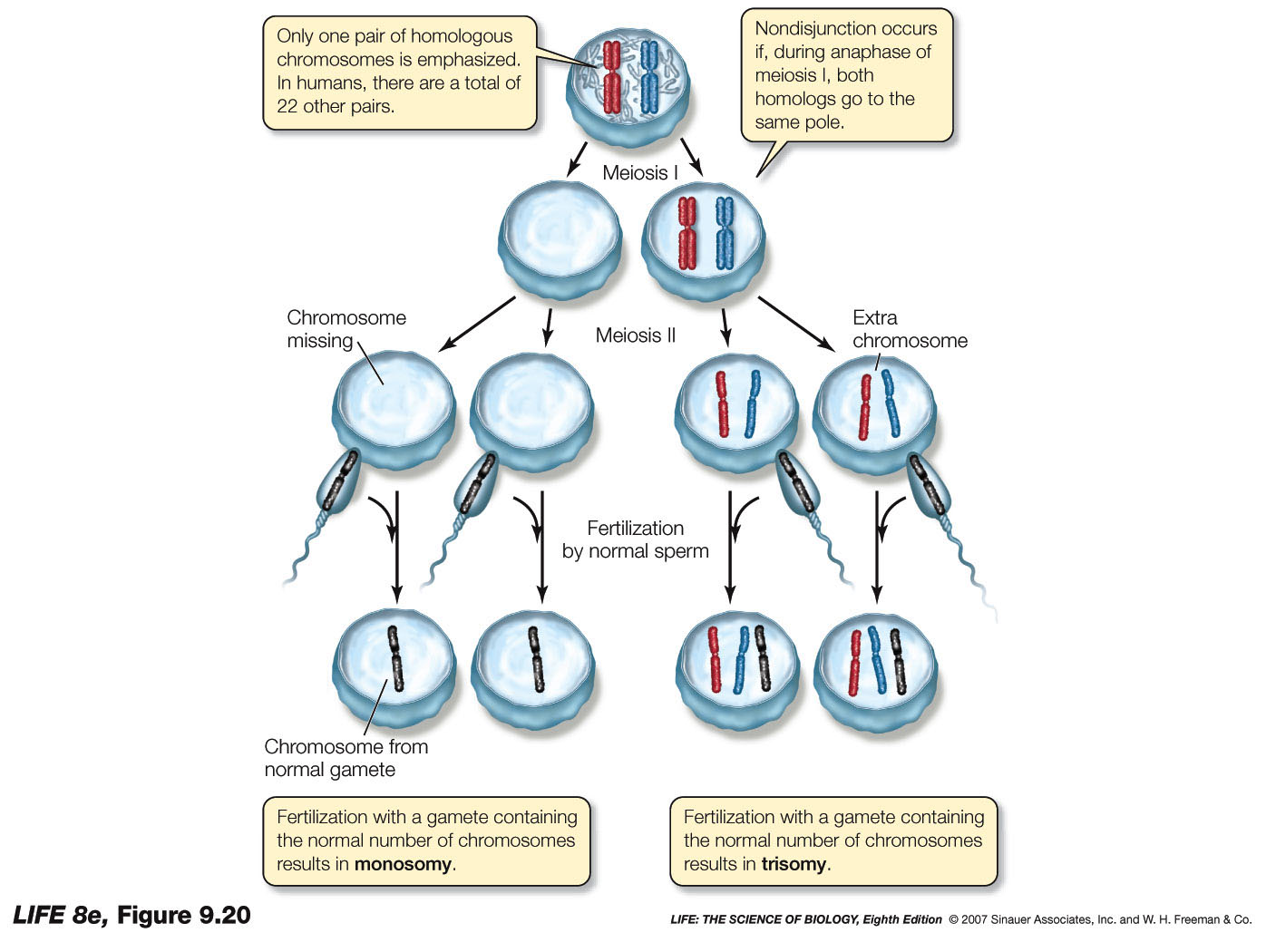
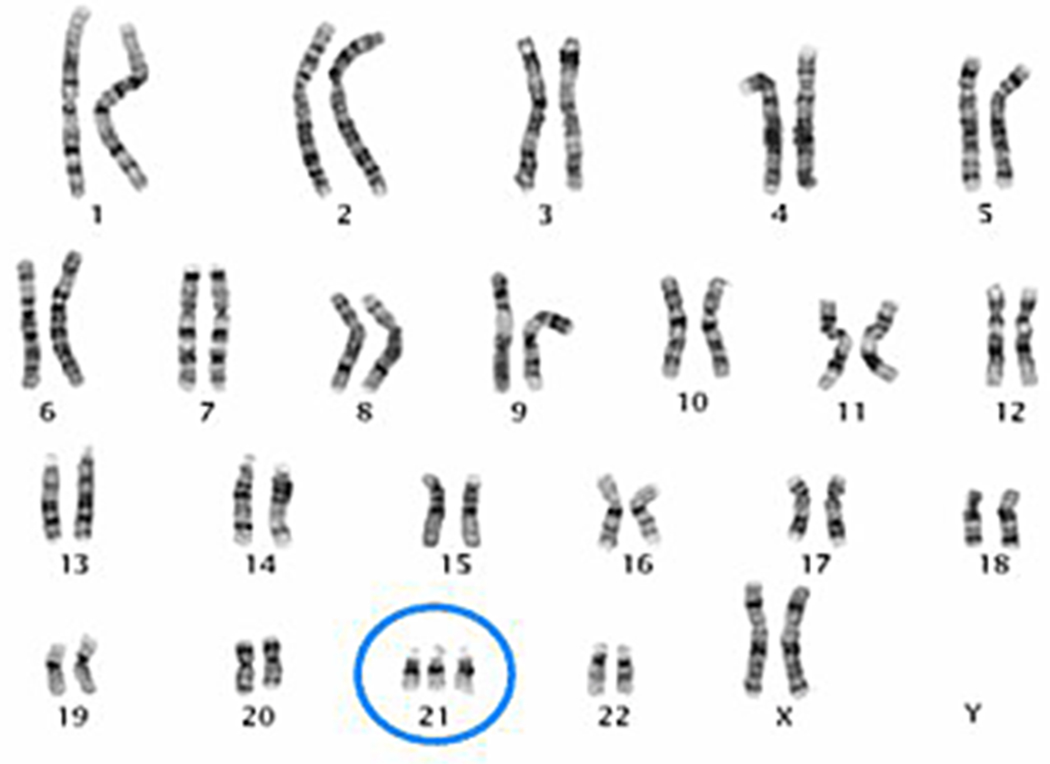
Aneuploidy, results, which can can give rise to a number of genetic abnormalities in the human condition, most of which do not survive past formation of the egg.
Meiosis demonstrates one of the apparent "truisms" of the living world, "all things biological come in pairs" (Francis Crick), where copies of the genetic information are transferred to the gametes by division of the germ-line cells into four "gametes" (two by two).Moreover, as we have seen, because of the packaging of DNA in anticipation of cellular division all of these features can be readily observed under the microscope.
Indeed, at the turn of the last century the whole process of the "dancing chromosomes" meiosis2.mp4 was correllated to the behavior of potential passage of "information" from one generation to the next Sutton and Boveri (1902).
At this time, while the microscope brought incredible insights in to how cells behaved, people's understanding of inheritance had not really changed from the 16th century idea of fully formed "homunculi" present in either the sperm of the father or somewhere within the mother... then, though some understanding of agriculture and obviously inheroted traites the idea of "blending" of parental traits came to the fore around the early 19th century.
Curiously, an Augustinian Friar by the name of Gregor Mendel had already finished and published his work almost forty years earlier (in 1865/1866), but no one had taken any notice of this unassuming Friar.
Even so, with 20/20 hindsight, we need not fall into the same trap and so can appreciate his accomplishments.
Mendel had already finished and published his work almost fourty years earlier (in 1865/1866) but no one had taken any notice of the unassuming Friar.
Mendel had many claims to fame, but one of his big contributions to science was that he devised a careful research plan, and used True-breeding plants.
Mendel’s well-organized research allowed him to "observe" and "record" the phenotypic traits of each generation in sufficient quantities to explain -through mathematical analysis- the relative proportions of the different "phenotypes" of the various progeny. Indeed, his paper is recognized today as a model of clarity.
He chose garden peas as his subjects, as they were easily grown and their pollination was easily controlled.
He controlled pollination by manually moving pollen between plants.

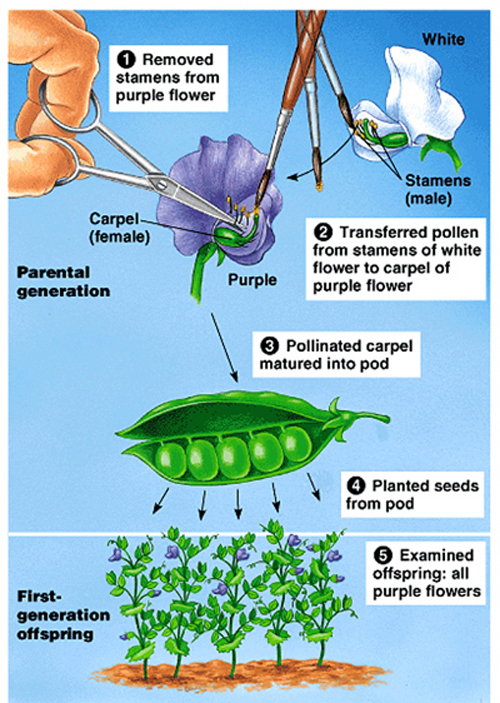
He
could also allow the plants to self-pollinate.
Mendel examined varieties of peas for heritable characters and traits
for his study.
A character is a feature, such as flower color.
He define a "trait" as being a particular form or a character, eg. white flowers.
Mendel looked for characters that had well-defined, alternative traits and that were true-breeding.

He considered a trait to be "true-breeding" when it was the only
trait that occured after many generations of breeding individuals that exhibited
that particular trait (we now call it phenotype).
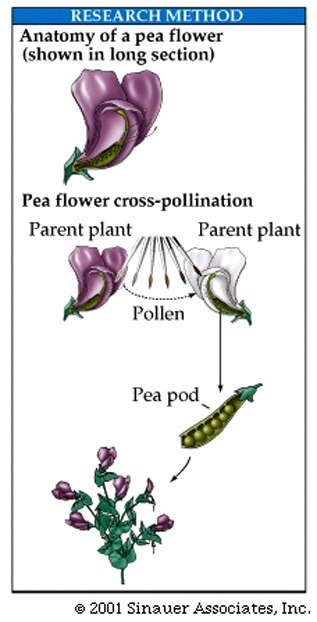
A true-breeding white-flowered plant would have only white-flowered progeny when crossed with others in its strain.
True-breeding plants, when used for crossing with other plants that have an alternative trait, arre called the parental generation, designated P.
The progeny from such a cross are called the first "filial generation", designated "F1".
When F1 individuals are themselves crossed to each other or "self-fertilized", their progeny are designated F2.
His first experiment examined what we now call a monohybrid cross
Mendel
crossed true-breeding plants that differed for a given character.
A monohybrid cross involves one (mono) character and different (hybrid)
traits.
Pollen from true-breeding pea plants with wrinkled seeds (one trait) was
placed on stigmas of true-breeding plants with spherical seeds (another
trait).

The F1 seeds were ALL spherical; somehow the wrinkled trait failed to appear at all.
Because the spherical trait appeared to completely MASK the wrinkled trait when two "true-breeding" parental plants are crossed, the spherical trait was called DOMINANT, and the wrinkled trait was called recessive.
But Mendeld didn's stop there. The F1 plants were then allowed to "self-pollinate".
This step defines the genius of Mendel, and is termed the monohybrid cross or an F1 cross.
The progeny, called F2, were examined: 5,474 were spherical and 1,850 were wrinkled.
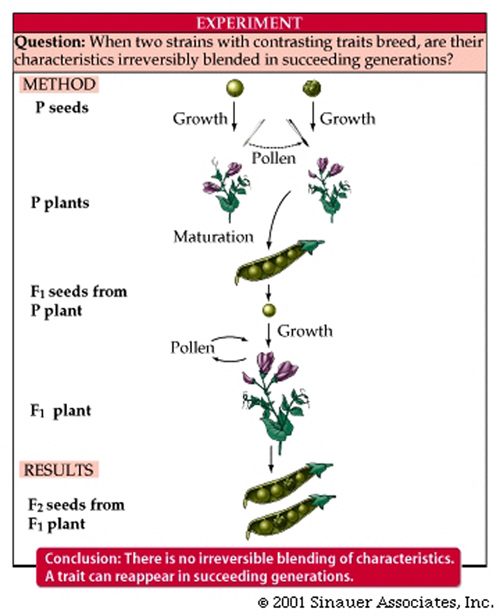
From his many experiments, Mendel proposed that the units responsible for inheritance were discrete particles, and that they existed within an organism in pairs; that they separated during gamete formation, and that they retained their integrity.
- this defined the particulate theory, which was in sharp contrast to the "blending theory", that was in vogue at the end of the 19th century.
Essentially, Mendel recogized that each pea had two units of inheritance that kept on showing through for each character.
Gregor Mendel's hypotheses:
1. Hereditary determinants are of a particulate nature. Each genetic trait is governed by unit factors , which "hang around" in pairs (or gene pairs) within individual organisms.
2. When two different unit factors governing the same phenotypical trait occur in the same organism, one of the factors is dominant over the other one, which is called the recessive trait.
3. During the formation of gametes the "paired" unit factors separate or segregate randomly so that each gamete receives either one or the other of the two traits, but only one .
4. The union of one gamete from each parent to form a resultant zygote is random with respect to that particular characteristic.
5. During production of gametes, only one of the "pair members" for a given character passes to the gamete.
6. When fertilization occurs, the zygote gets one from each parent, thus restoring the pair.
You can see where we are going with this, but at the time........
Mendel’s
units of inheritance are now equivalent
to the genetic units or genes.
Different forms of which are called alleles.
Each allele is given a symbol. In the case of wrinkled seeds, "S" might represent smooth and "s" wrinkled.
By a subsequent convention, uppercase always represents the dominant trait (if it is known), while the lowercase represents the recessive.
If Mendel's appreciation of heredtary holds true then "true-breeding" individuals would each have two copies of the same allele.
Wrinkled
would be ss. (two copies of same
allele = homozygous)
Smooth true-breeding
would be SS. (two copies of same
allele = homozygous)
However, if this were to be the case, as a consequence of a cross between true-breeding round and true-breeding wrinkled seed plants, the smooth-seeded plant offspring would be Ss, and would not themselves be true-breeding parents for their own offspring, and would be considered to be heterozygous.
The actual composition of any organism's alleles for a gene is its genotype: Ss would, therefore, be a genotype.
When an individual produces gametes, its alleles separate, so that each gamete receives one member of the pair of alleles.
Giving rise to Mendel's first law, the law of segregation.
| Mendel's First Law: Two members of a gene pair segregate from each other into the gametes, whereby one half of the gametes carries one of the traits, the other half carries the other. |
To analyze how this might come about at the turn of the last century a Cambridge Professor (William Punnett) devised, what is now considered to be the Punnett square, which is a simple box-like device that helps us to consider all genetic combinations and can provide clarity by showing the expected frequencies of genotypes.
The Punnett square shows that the genotypes and associated ratios for a monohybrid cross are:1 SS : 2 Ss : 1 ss. even though the "phenotype" of the progeny exhibits a phenotypic ratio of (approximately) 3 smooth to 1 wrinkled.
The 2nd Punnett square shows a similar set of genotypes and the associated ratios for a monohybrid cross of these traits are:1 YY : 2 Yy : 1 yy. Again, even though the "phenotype" of the progeny exhibits a phenotypic ratio of (approximately) 3 yellow to 1 green.
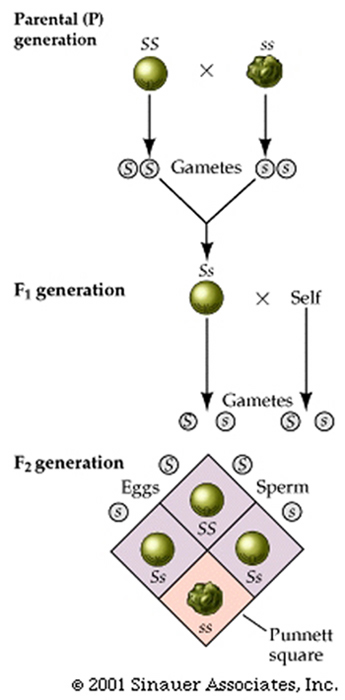
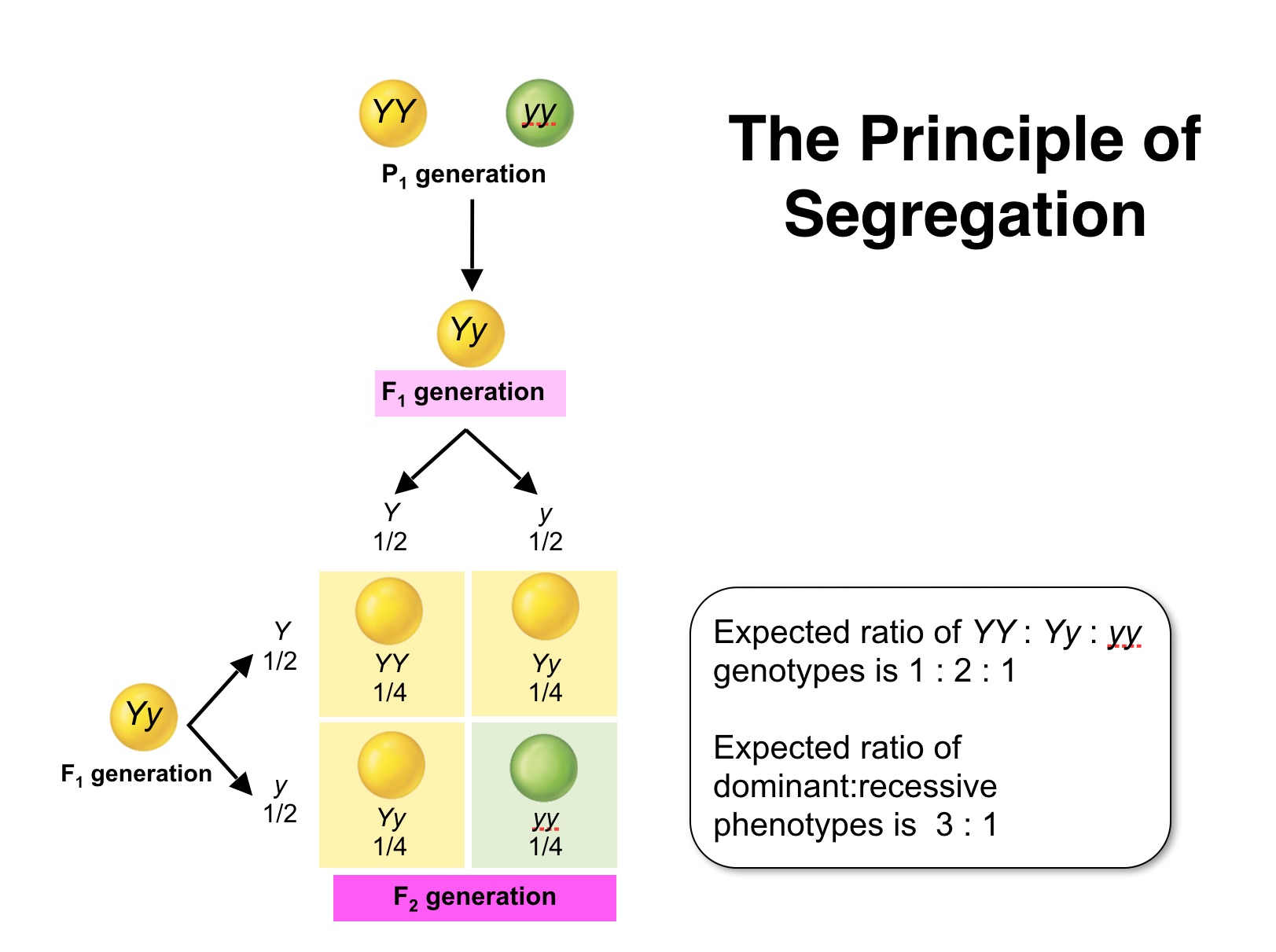
Indeed, ALL the traits that Mendel analyzed appeaared to have the same 3: 1 ratios in the F2 generation of crosses of plants that were "pure bread" yielded a singular "dominant" trait in the F1 generation .

Mendel verified his hypothesis by performing what is now termed a test cross, which can determine the genotype (heterozygous or homozygous) of an individual that may exhibit a dominant phenotype of a given trait.
It involves crossing the "unknown" individual to a true-breeding recessive or homozygous recessive.
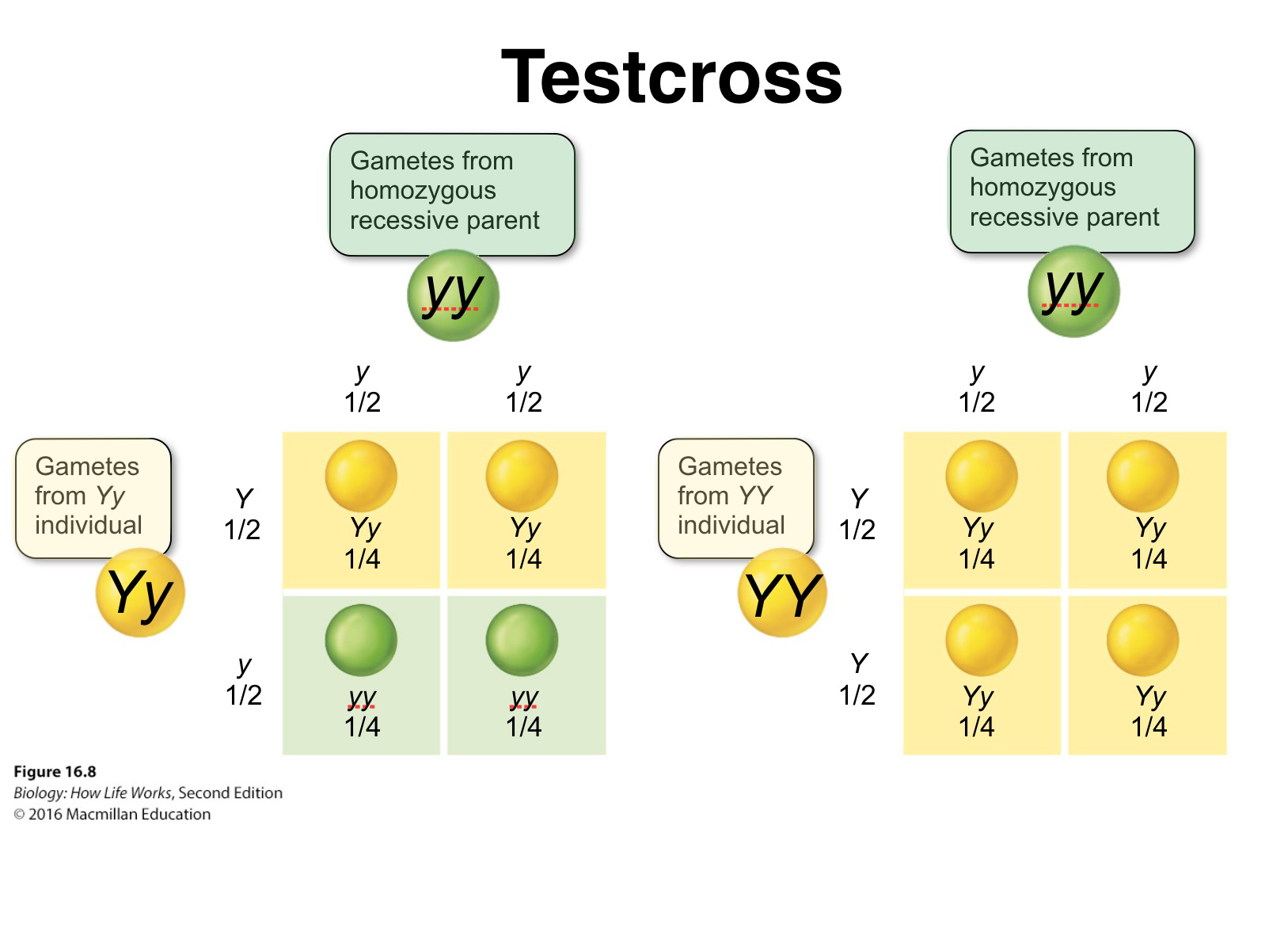
If the unknown is homozygous dominant for the particulat trait what is the outcome?
If the unknown is heterozygous for a given trait, approximately half the progeny will have the dominant trait and half the recessive trait.
The Punnett square shows that the genotypes and associated ratios for a monohybrid cross are:1 SS : 2 Ss : 1 ss. even though the "phenotype" of the progeny exhibits a phenotypic ratio of (approximately) 3 smooth to 1 wrinkled.
While it is now known that a gene is a portion of the chromosome that resides at a particular site, called a locus (plural being loci).
Remember, Mendel had no knowledge of this or meiosis, which nicely explains his law of segregation. (See Animation Movie)
Mendel verified his hypothesis by performing what is now termed a test cross, which can determine the genotype (heterozygous or homozygous) of an individual that may exhibit a dominant phenotype of a given trait.
It involves crossing the "unknown" individual to a true-breeding recessive or homozygous recessive.
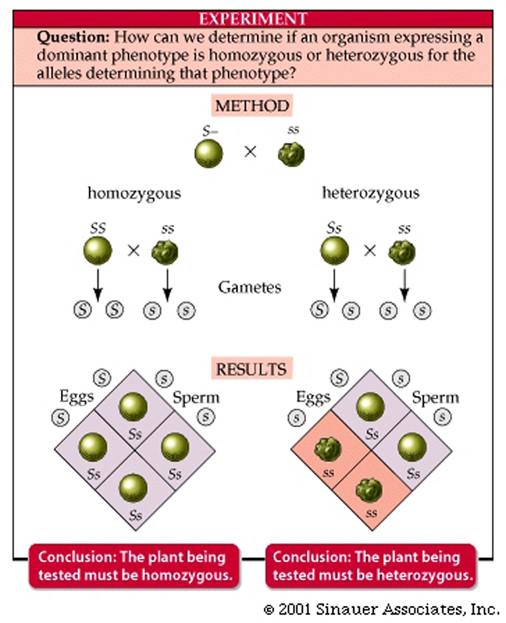
If the unknown is homozygous dominant for the particulat trait what is the outcome?
If the unknown is heterozygous for a given trait, approximately half the progeny will have the dominant trait and half the recessive trait.
Mendel's second law is the law of random assortment and simply states:
Mendel's Second Law: During gamete formation the segregation of one gene pair is independent of all other gene pairs |
This second law describes the outcome of
a dihybrid (two character) cross, or hybrid cross involving additional
characters/traits.
A
dihybrid is an individual that is a double
heterozygote (e.g., with the genotype SsYy).
Mendel’s second law states that the Ss alleles assort into
gametes completely independent of the Yy alleles.
Thus, the dihybrid, Ss Yy, produces gametes that have one allele of each gene.
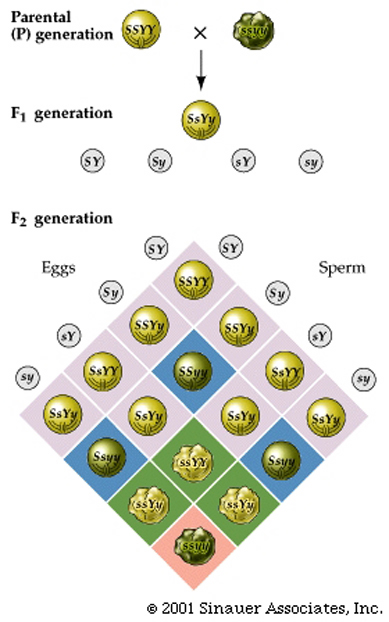
In this cross, Four different gametes are possible and will be produced in equal proportions: SY, Sy, sY, and sy.
Random fertilization of gametes yields the outcome visible in the more complex Punnett square.
Note that it is now 4 x 4 table construction to accommodate 16 possible genotypes.
Filling in the table and adding the like cells reveals a 9:3:3:1 ratio of the four possible phenotypes (smooth yellow, smooth green, wrinkled yellow, and wrinkled green).
Monohybrid crosses, dihybrid crosses -and even more complex- trihybrid crosses: same principles.

Know how to test for Mendel's laws.
While Mendel could not comprehend any of the genetic intricaies that his laws might suggest, his determination of his abstract "unitary" traits had far reaching implications in understanding inheritance of these genetic traits and how they could be inherited.
As we have discussed, each of the "gene variants" that Mendel called "factors" are termed Alleles and ALL variants of a given gene reside at a specific locus on a given chromosome.
Variation in allelic types arise by mutation or change in the DNA sequence.
An allele,
therefore, can arise due to a mutation
at a given locus, and can be analyzed through classical genetics once that variation has been incorporated
Alleles can be randomly mutatagenized
to become a different allele, depending
on DNA sequence changes and maintained or lost -again in a random manner- unless
acted upon by a selective pressure.
"Wild Type", as we have discussed prevously, is a term used to demonstrate the most common allele/trait of a given population. Again, being the "most common" allele in a population does not always mean that the wild-type allele will be the dominant or "superior" allele.
However, some definition of "normalcy" in the genetic expression of traits does provide an incredible advantage in the understanding of how genes work in the whole organism...
In the early twentieth century, G. H. Shull crossed two varieties of corn, and the yield went from 20 to 80 bushels per acre, thus defining a, now common, agricultural practice to increase production in plants.
{kind=link}

This is called either hybrid vigor or heterosis, whereby crossbred strains show superior qualities, and it is a result of the fact that continued genetic "in-breeding" often leads to deleterious traits becoming "fixed" in the gene pool (an aspect we wil address later). The mixing of two strains, therefore, reintroduces a number of selectible traits which can enhance the gene pool of the newly defined strains... Use of such hybridization techniques has been a common agricultural practice ever since -to increase production in crops and broadened to cattle etc...
An hypothesis called overdominance even proposes that the heterozygous condition -in certain important genes- is often considered to be selectively "superior" or have a higher fitness than the presence of either homozygote. This occurs far more frequently in cases where a single trait controls more than one phenotype. Such heterosis, however, would be slightly at odds with Mendel's idea of "independent" traits.
Even so, given the potential of heterosis (cross bred alleles show some qualities superior to their parental counterparts), this would threaten the very crux of Mendel's ideas which are premised on the "particulate" nature of genes, and where one allele in any given gene pair is simply dominant over the other
Perhaps, Mendel wasn't perfect...
No, he wasn't, he was also only observing, NOT deducing. Even so, the fact that we are still talking about him is testament to how much he did get right.

About Contact GSU Academics Degrees & Majors |
Admissions Undergraduate Research Dept. of Biology |
Libraries University Library Campus Life Housing |
Athletics Alumni
|