

- Home
Welcome !
Welcome to john houghton's home page for his biology courses. This site is designed as a hub for curating and sharing lectures, course syllabi, assignments, and links to relevant resources. Use the menu bar at the top of the screen to navigate through the site.
(Please note: this page is currently under construction.)

- BIOL 2107
Fall '23 CRN86772
Lectures: (1)
- Courses
BIOL 2107 Principles of Biology I

| Lecture 26: Course Overview and Introduction to the "Meaning" of LIFE |
Central Dogma 3D -Animation Central Dogma REAL-TIME -Animation
Prokaryote Regulation I Prokaryote Regulation II Eukaryote Regulation
Having dealt with many aspects of the Central dogma, as highlighted by the two animations above, it is important to try to bring EVERYTHING that you have learned in the last nine lectures or so lectures into a context that will allow you to discuss more general questions.
Yes, for the purpose of doing well on the Final, but principally because this is the only way that you will more fully understand the concepts that have been discussed in the lecture series.
This lecture series is also supposed to begin to allow you to use what you have just learned, and to so effectively; giving you some glimpse as to some of the complexities that are present in all biological systems... as a springboard for future lecture series in Biology.

To emphasize this point, this lecture is going to begin to discuss some of the complexities of regulation in the central dogma: higher orders of control and how they are similar in prokaryotes and equkaryotesand yet differ substantially in these to domains of living organisms.
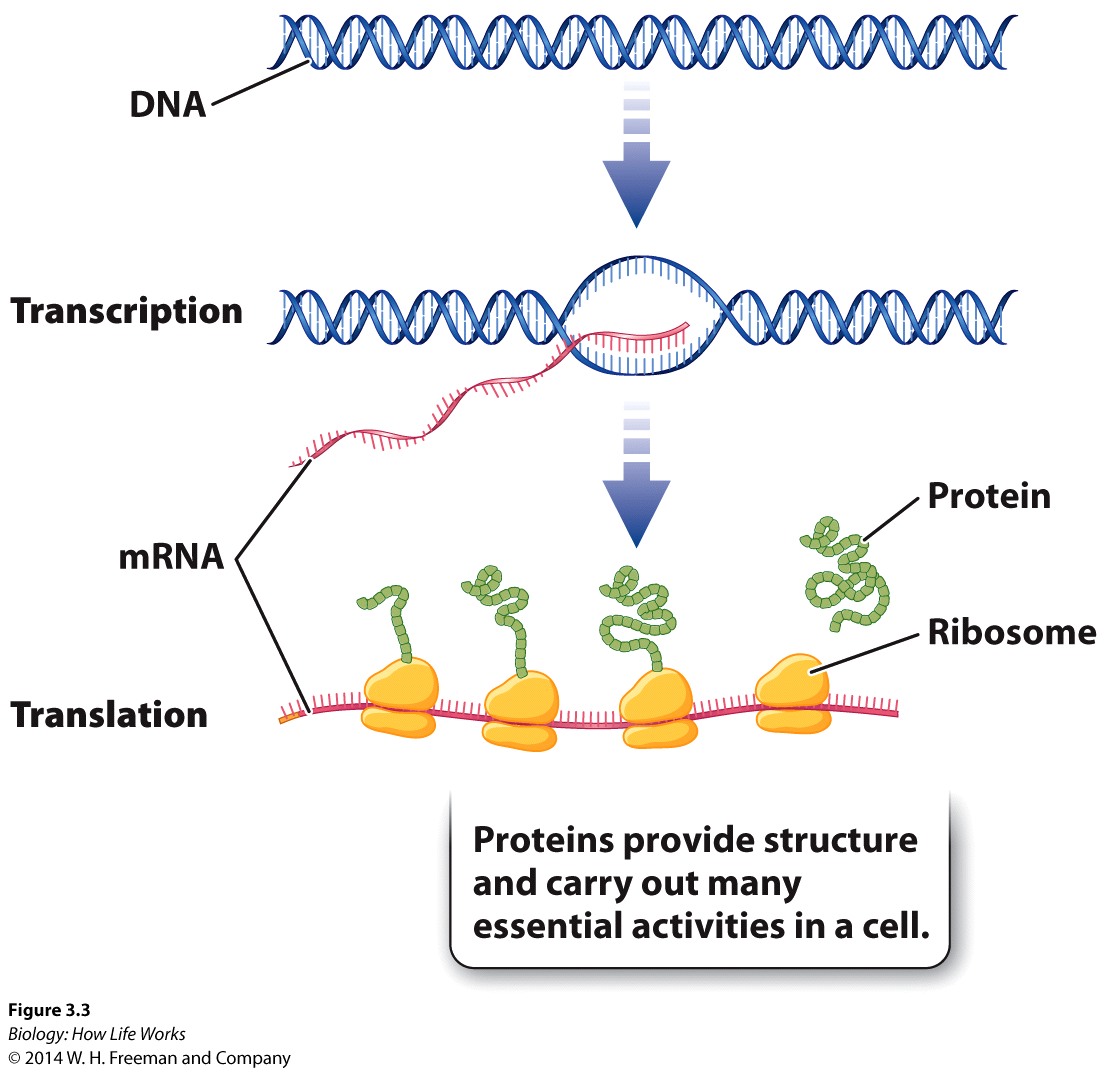

While regulation occurs at every level of the central dogma for each of the myriad numbers of genes in any organism, it is important -in order for us just to get a "hint" of the complexity of control within living organisms to appreciate regulation at just one level and in the simplest system, the bacterium, E. coli.
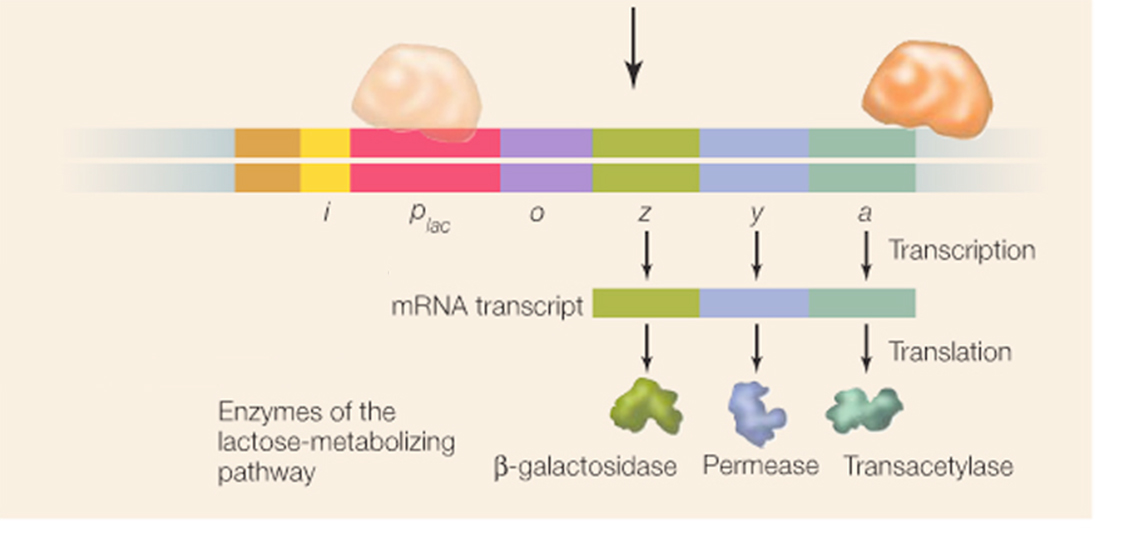
The first level of control can be understood by understanding how genes involved in the break down of an important secondary sugar called lactose is controlled in bacterial systems (such as E. coli). This bacterium controls expression the three genes that give rise to three enzymes involved in the breakdown of this sugar by putting all three genes behind a single promoter. In so doing it controls transcription of all the associated genes that have been clustered together in the same region of the chromosome. Their synthesis, therefore, is said to be coordinated within a sequence of genes called an "operon"
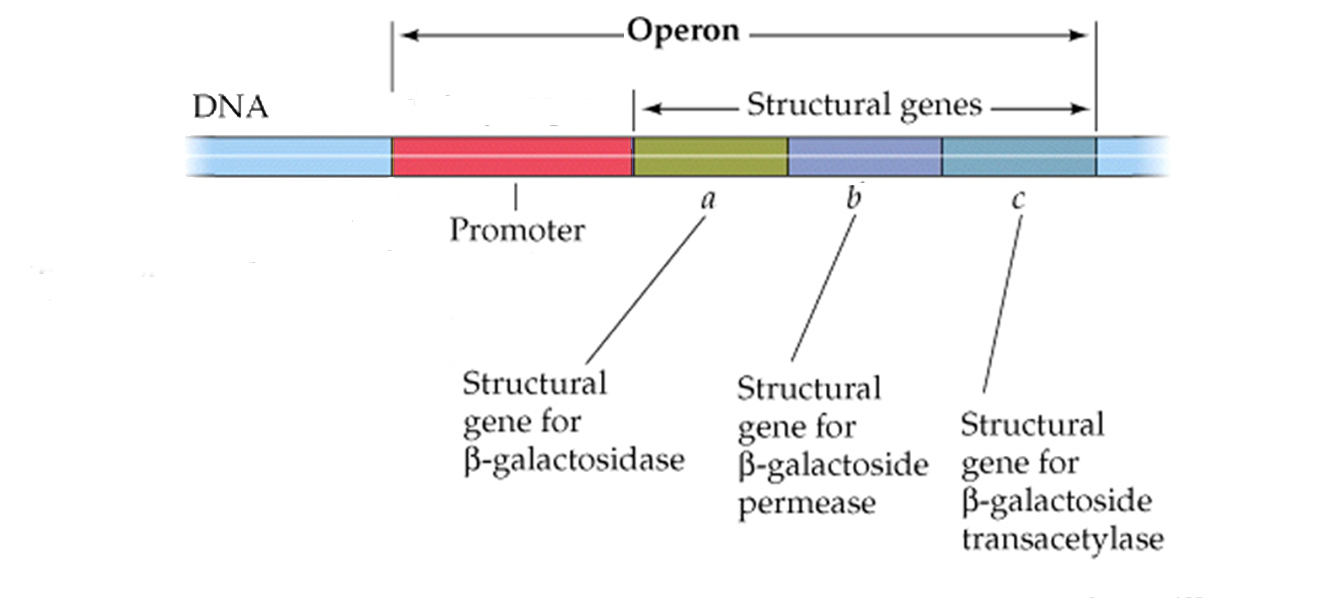
Complete shut-down of transcription in prokaryotes can occur by placing an obstacle between the promoter and its structural genes; by introducing a new section of DNA called the operator.
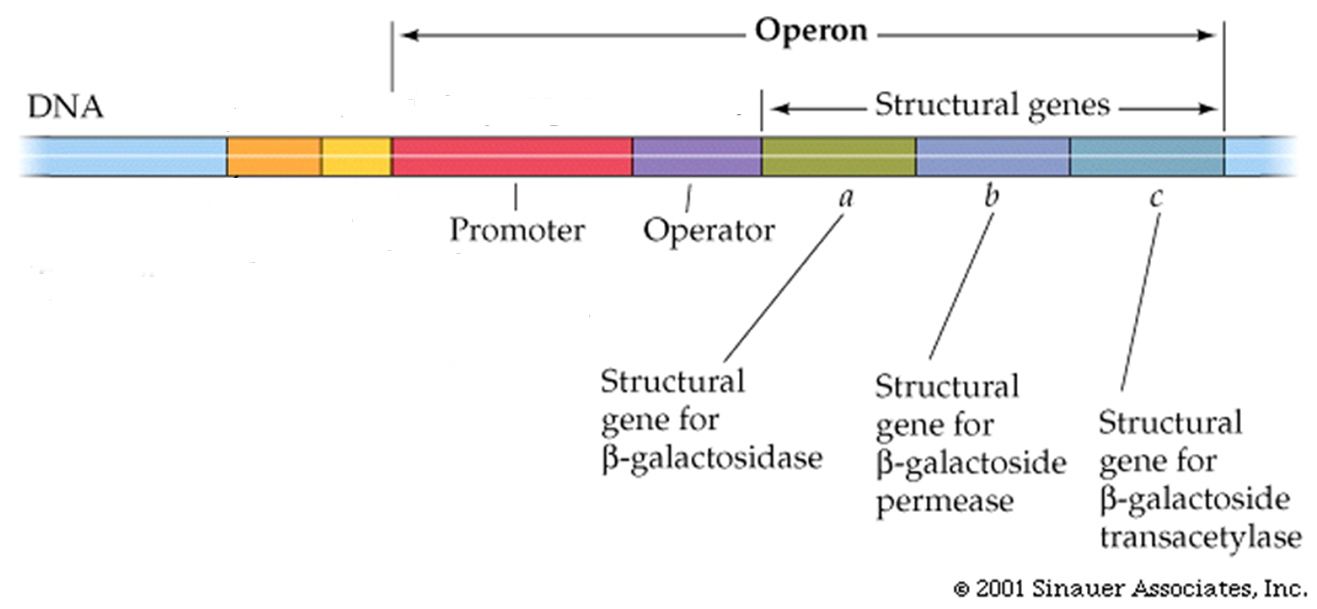
If a specific protein, the"repressor",is able to bind to the operator, it creates an obstacle, binding and/or movement of the RNA polymerase is blocked from transcribing the structural genes.
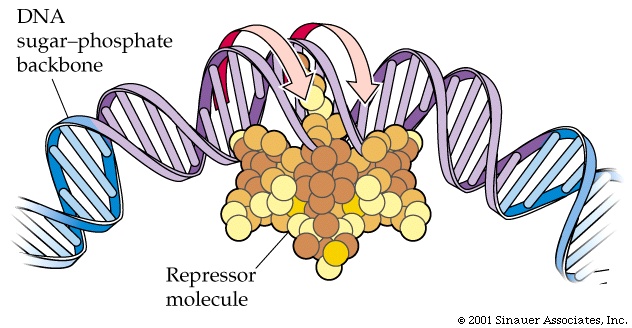
But now the bacterium has to synthesize a gene that makes this specific repressor protein, so why not put that gene in the same area of the genome, close to the operon it's controlling.
But all this does is provide a system that is always "Turned OFF" as the repressor protein ALWAYS binds the "operator" and blocks transcription.
So, to make this system regulatable, you will need to modify the repressor protein which blocks transcription to be able to bind the operator most of the time, but sometimes come off the DNA and allw transcription to occur. This repressor protein, would need to be an allosteric protein and have TWO binding sites: one for the operator and the other for the inducer, which changes the shape of the repressor protein and disallows its binding to the operator.
This is exactly what happens in prokaryotes.
Example:
We have looked at catabolism of sugars, and how -like ourselves- a lot of organisms preferentially thrive on a Glucose diet. E. coli, is one such bacterium that prefers glucose, but when glucose availability is low and other sugars - such as lactose- are in abundance, E. coli can use these other sugars... in this example lactose.
Lactose is a disaccharide containing galactose - linked to glucose.
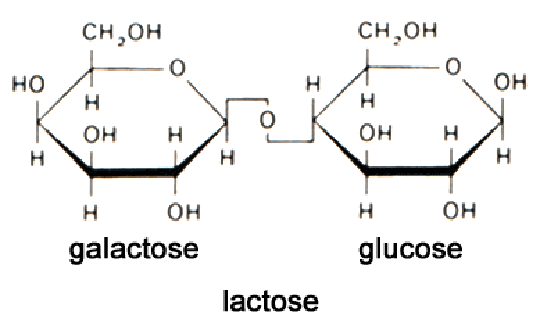
Lactose is transported into the cell by a carrier protein (enzyme) called ß-galactoside permease, whereupon the lactose is hydrolyzed to glucose and galactose by the enzyme ß-galactosidase.
Another enzyme is needed to protect the cell from the side effects of too much ß-galactosidase enzyme activity. It is called thiogalactoside transacetylase.
So, because all these genes are involved in the same process (metabolism of lactose), all these genes are clustered together in the genome and are synthesized on the same RNA transcript in what is called the lac operon.
To conserve energy, not too surprisingly, when no lactose is present, levels of all three enzymes should be and are low.
It turns out that the molecule, Lactose itself is able to activate (induce) expression of the operon and express large amounts of all three enzymes.
Remember that repressor protein....
The repressor protein fro the lac operon is called the lac repressor, and is coded for by the regulatory gene, lacI.
The lacI gene just happens to be located near the lac structural genes. However, not all regulatory genes are near the operons that they control.
Regulatory genes like lacI have their own promoters. In this instance, the lacI gene is expressed constitutively (i.e., it is expressed is all the time).

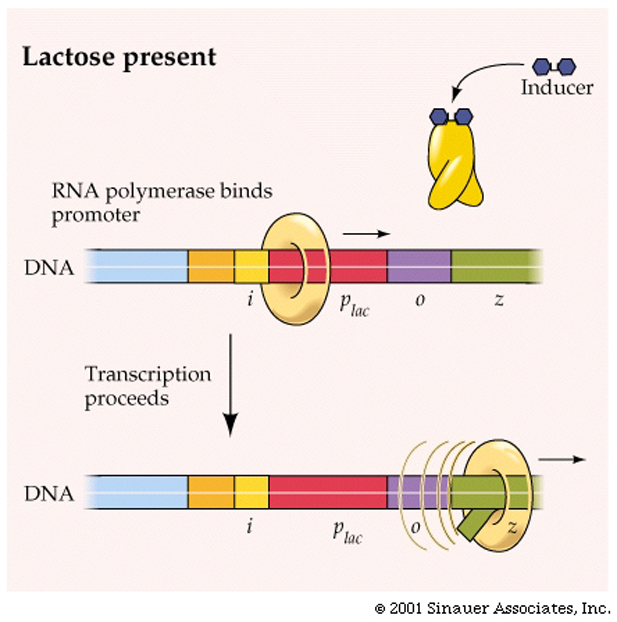
This form of regulation is one of the more simple examples of gene regulation, but it does highlight the importance of "coordinated" control of gene expression in controling the expression of genes - a vitally important aspect of regulating the Central dogma, and our understanding of how life really can, and does, exist.
So, what about Eukaryotes.......?
In eukaryotes the first level of organization of the genes is more simplified than organization of genes in prokaryotes. Most genes are "monocistronic" i.e. one mRNA (mature mRNA) molecule gives rise to only one gene product.
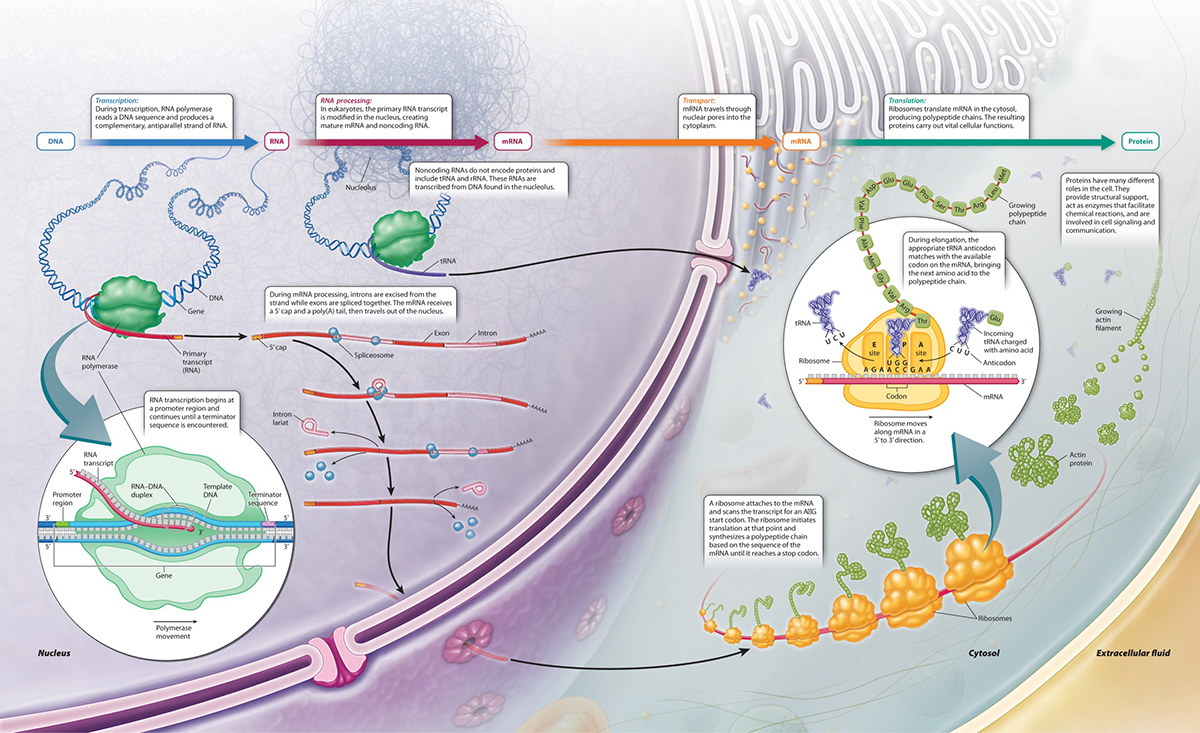
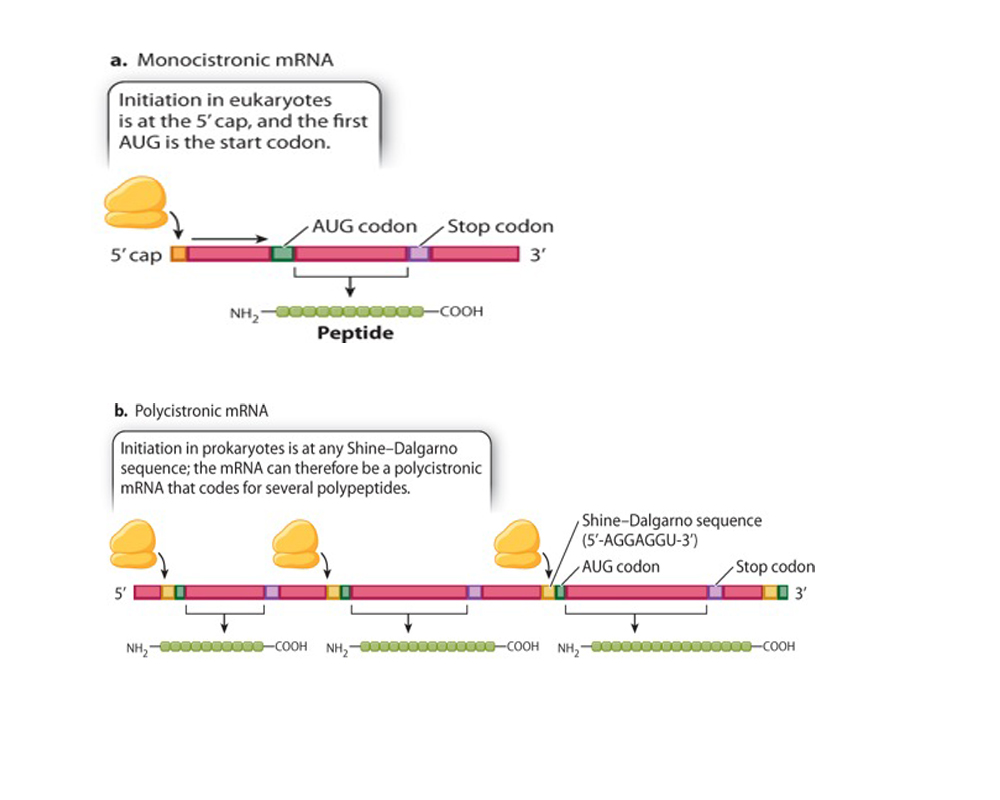
Eukaryotes also utilize a a similar system of competiion of repressors vs. activating proteins......theyalso try to solve different problems and have a lot more complexity, which requires considerably higher orders of regulation for the transciptional process itself.....
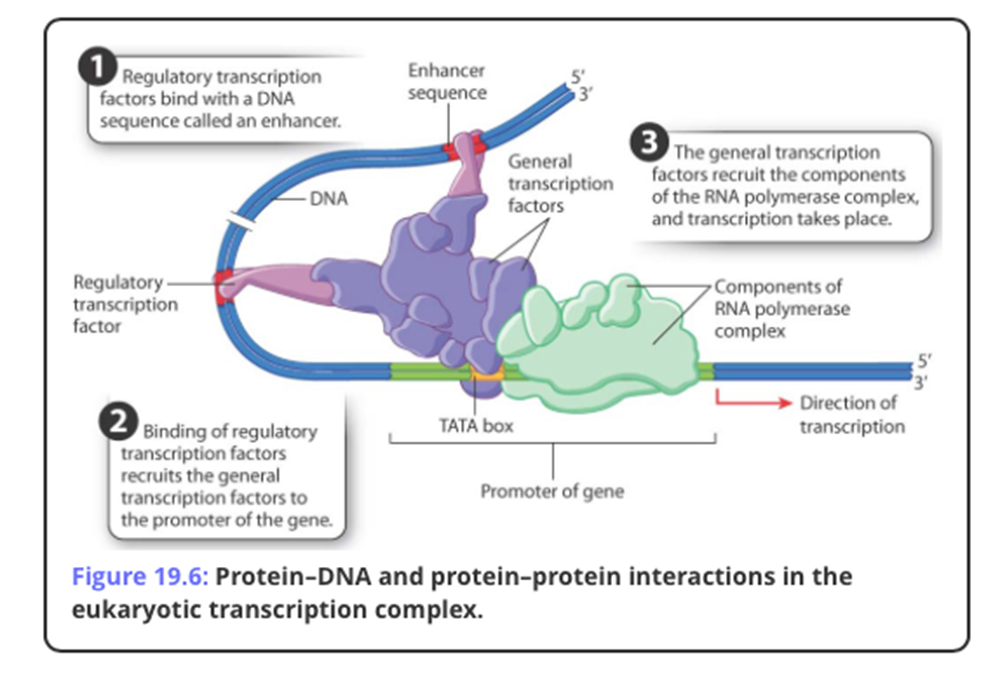
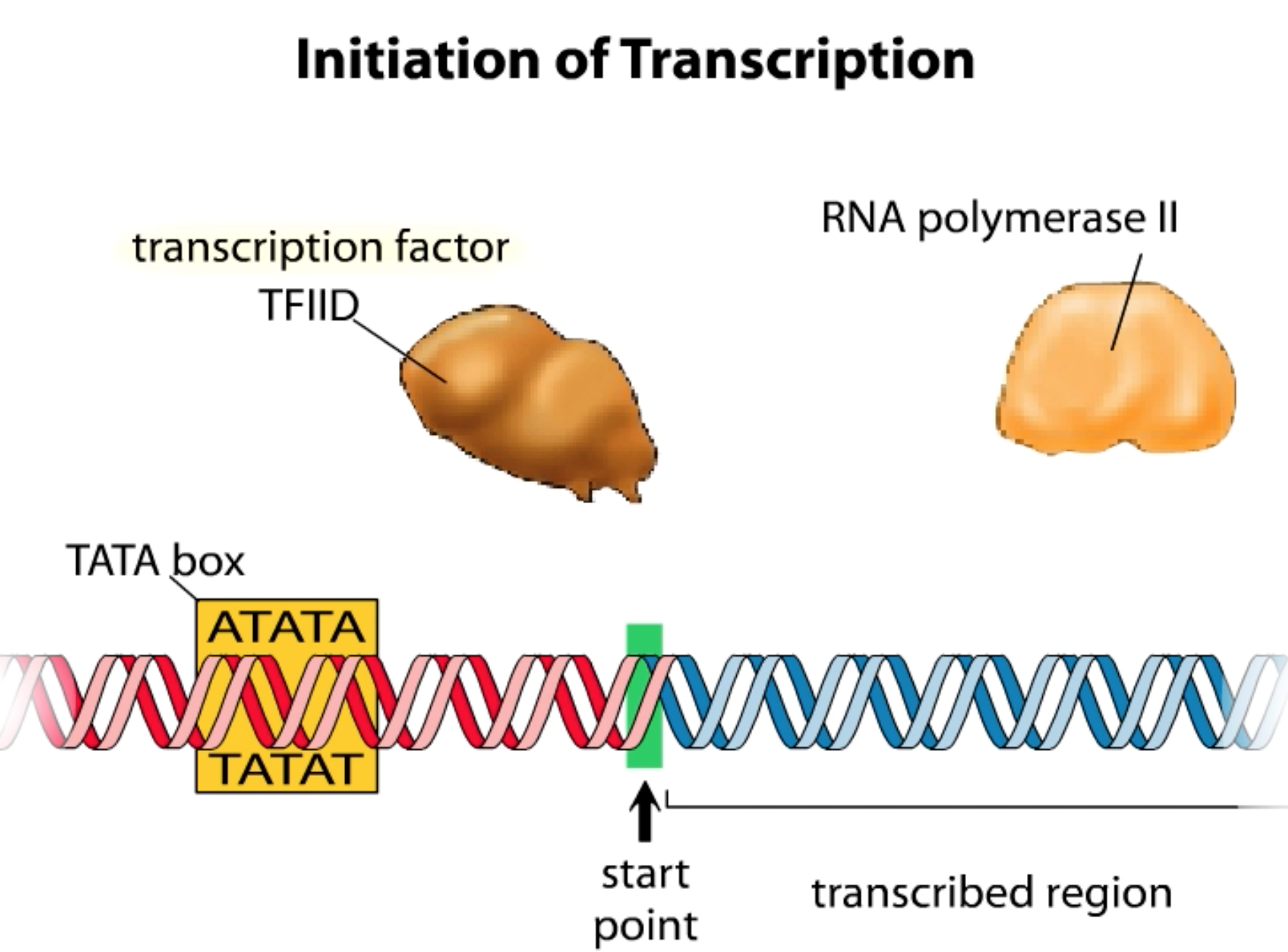
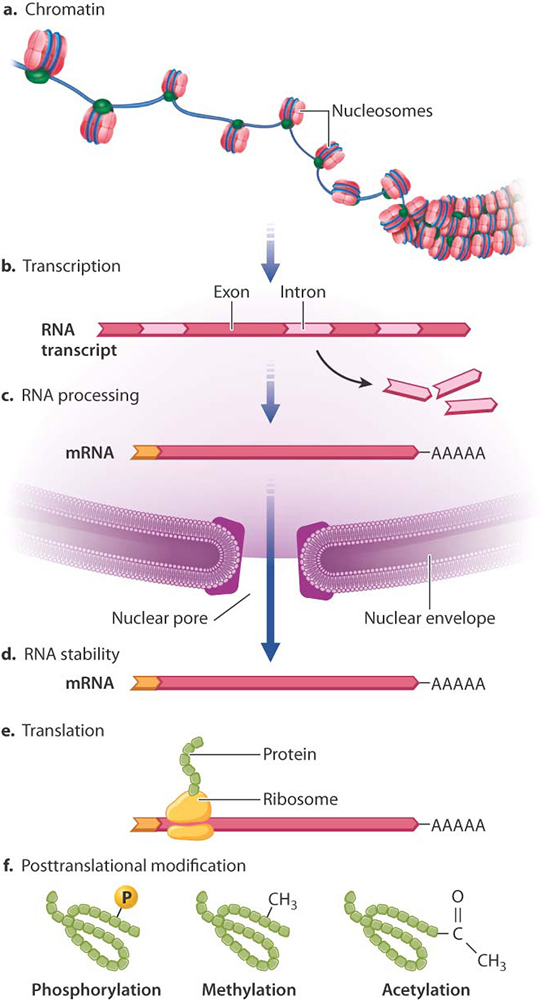
While there are always controls for each of the different steps in the central dogma....these are but the first steps In eukaryotes there are also a number of other layers of regulation that affect/ control gene expression. Some of these are NOT even defined in the DNA and, as such, are considered to be BEYOND genetic events or Epigenetic: none genetic influences on gene expresssion.
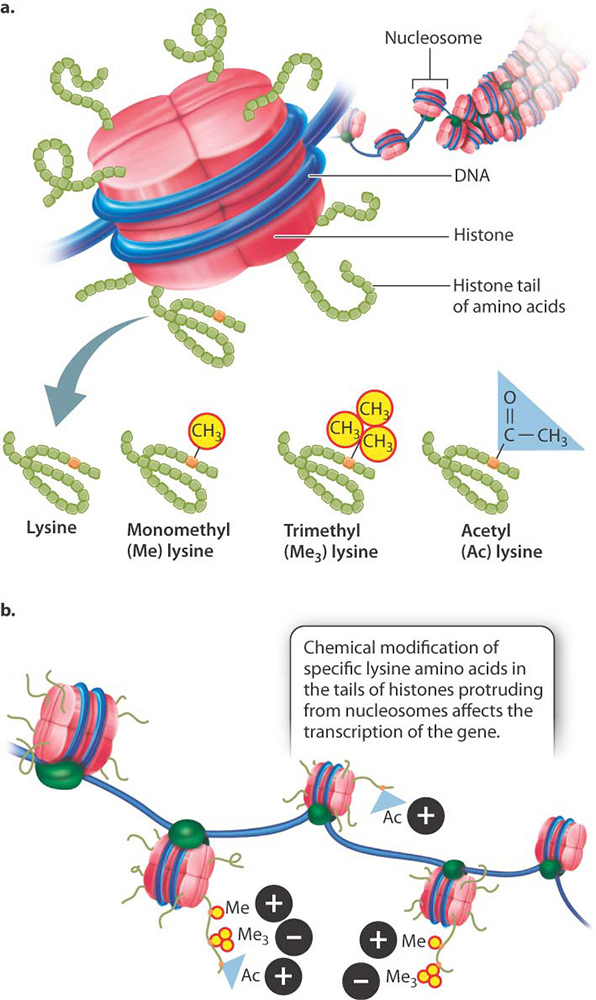
At various times in the lecture series, we have discussed the fact that, in eukaryotes, the DNA is present as, not juts as DNA, but as DNA wrapped around some proteins called "histones", collectively know as chromatin.
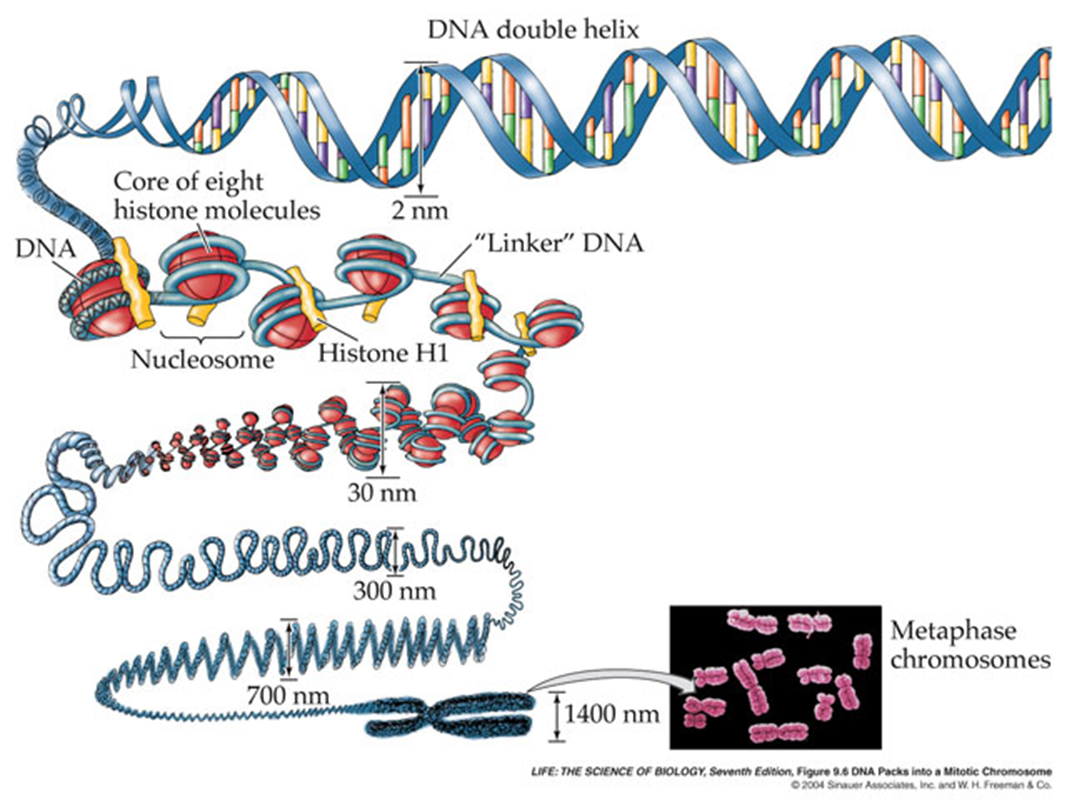
This packaging of the DNA, wrapped around these nucleosomes, is highly flexible and the density of this packaging can have a dramatic affect upon expression of the genes within the DNA itself.
The precise details as to how this works and, indeed, the precise mechanism(s) as to how global control of gene expression in different organisms is determined is not part of this course, but adds a level of intrigue (I hope) for other courses in Biology, yet to come.


About Contact GSU Academics Degrees & Majors |
Admissions Undergraduate Research Dept. of Biology |
Libraries University Library Campus Life Housing |
Athletics Alumni
|