

- Home
Welcome !
Welcome to john houghton's home page for his biology courses. This site is designed as a hub for curating and sharing lectures, course syllabi, assignments, and links to relevant resources. Use the menu bar at the top of the screen to navigate through the site.
(Please note: this page is currently under construction.)

- BIOL 2107
Fall '23 CRN86772
Lectures: (1)
- Courses
BIOL 2107 Principles of Biology I

| Lecture 12: Mendelian Genetics II |
A Scientific Law is "a descriptive generalization about how some aspect of the natural world behaves.... under stated circumstances".
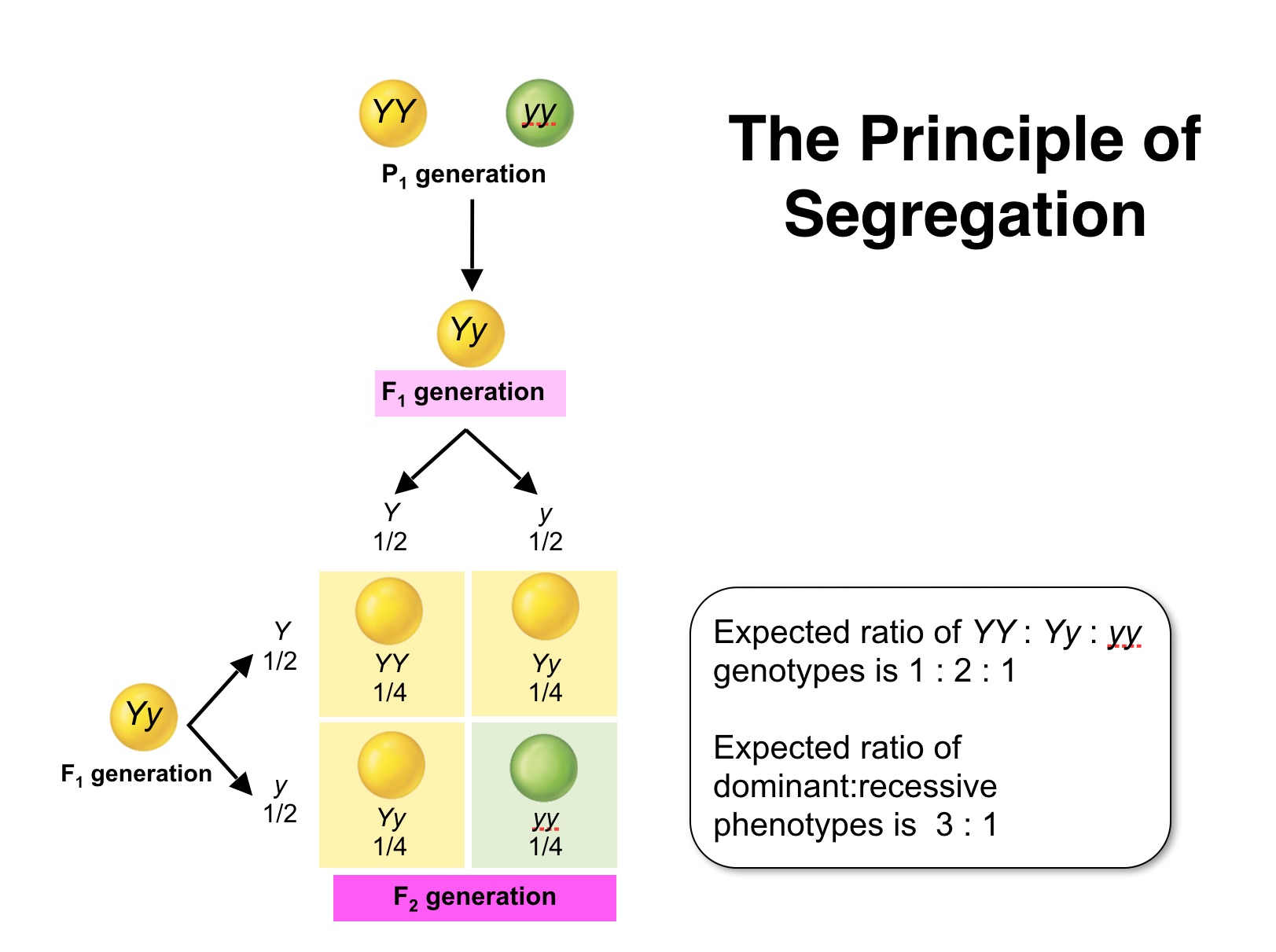
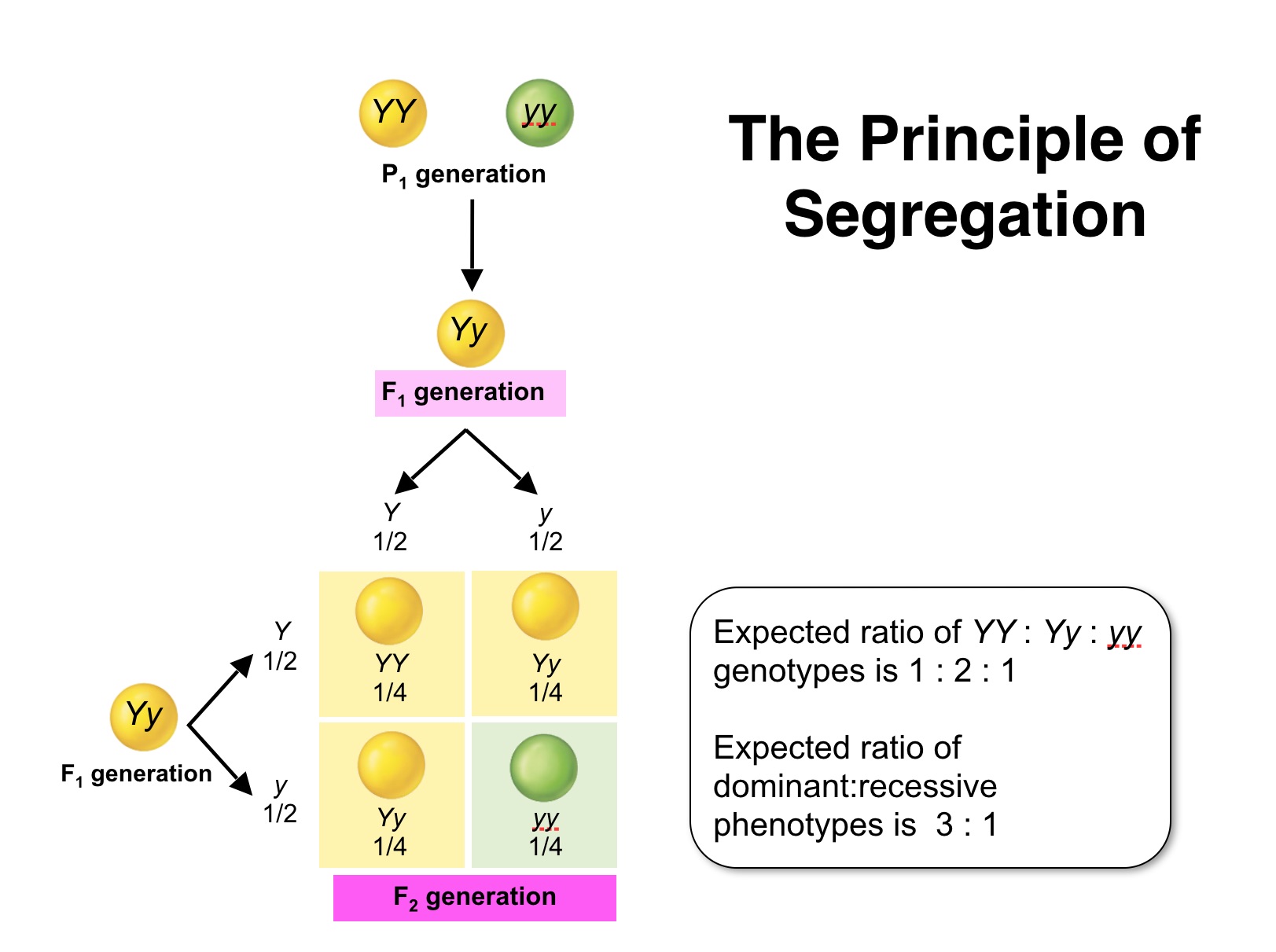
| Mendel's First Law: Two members of a gene pair segregate from each other into the gametes, whereby one half of the gametes carries one of the traits, the other half carries the other. |
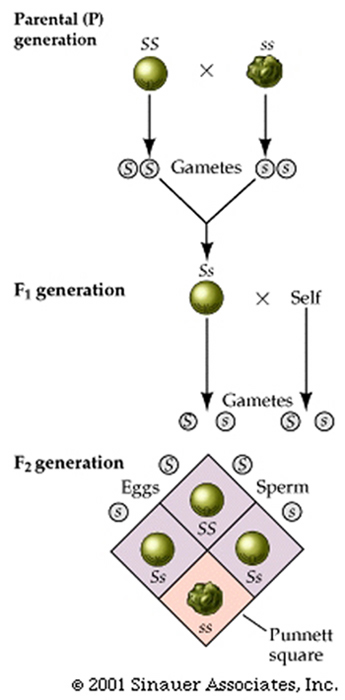
To analyze how this might come about a turn of the last century, Cambridge Professor (William Punnett) devised, what is now considered to be the Punnett square, which is a simple box-like device that helps us to consider all genetic combinations and can provide clarity by showing the expected frequencies of genotypes.
The Punnett square shows that the genotypes and associated ratios for a monohybrid cross are:1 SS : 2 Ss : 1 ss. even though the "phenotype" of the progeny exhibits a phenotypic ratio of (approximately) 3 smooth to 1 wrinkled.
While it is now known that a gene is a portion of the chromosome that resides at a particular site, called a locus (plural being loci).
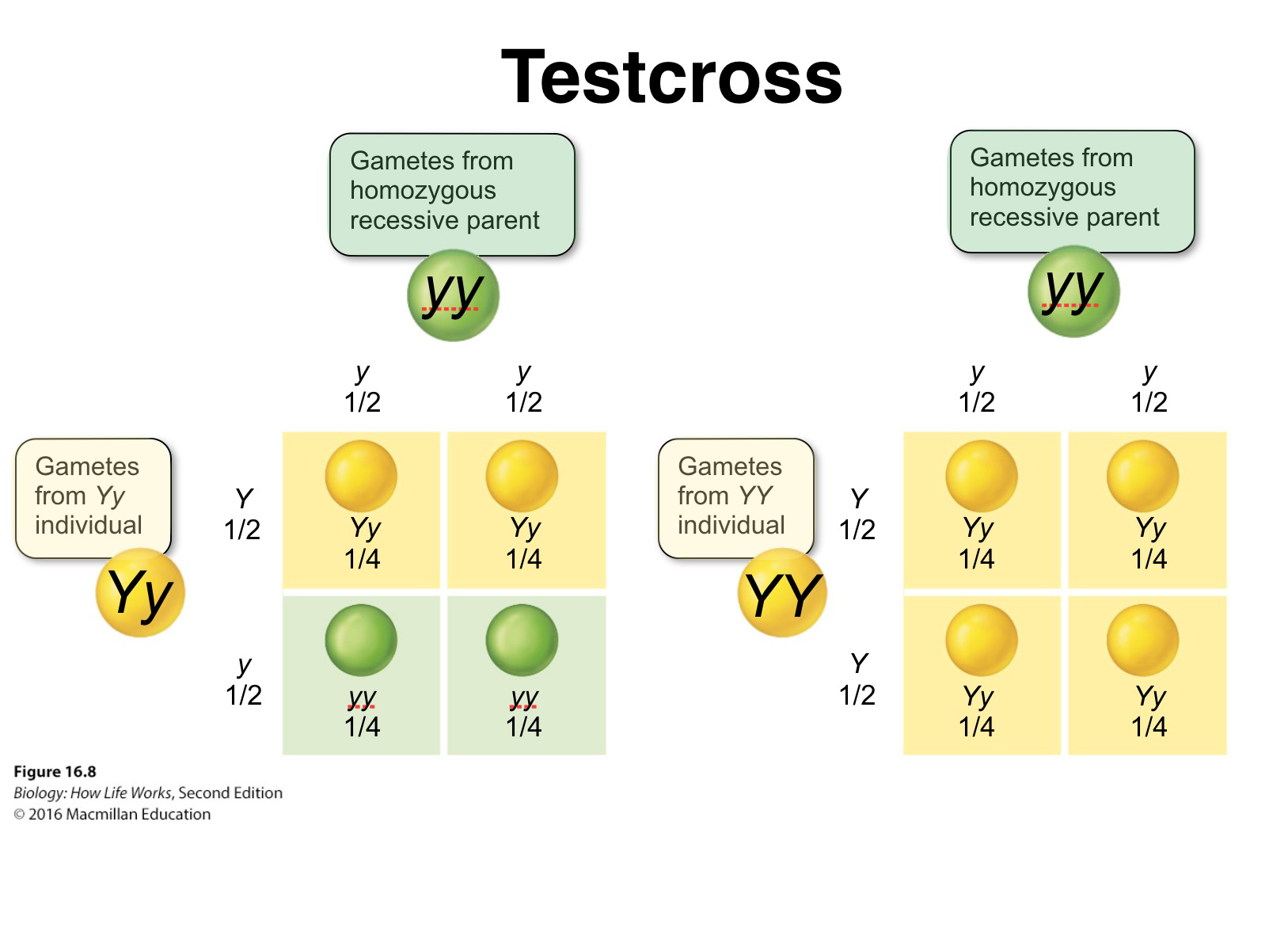
Mendel verified his hypothesis by performing what is now termed a test cross, which can determine the genotype (heterozygous or homozygous) of an individual that may exhibit a dominant phenotype of a given trait.
It involves crossing the "unknown" individual to a true-breeding recessive or homozygous recessive.
If the unknown is homozygous dominant for the particulat trait what is the outcome?
If the unknown is heterozygous for a given trait, approximately half the progeny will have the dominant trait and half the recessive trait.
Mendel's Second Law: During gamete formation the segregation of one gene pair is independent of all other gene pairs |
This second law describes the outcome of a dihybrid (two character) cross, or hybrid cross involving additional characters/traits.
A
dihybrid is an individual that is a double
heterozygote (e.g., with the genotype SsYy).
Mendel’s second law states that the Ss alleles assort into
gametes completely independent of the Yy alleles.
Thus, the dihybrid, Ss Yy, produces gametes that have one allele of each gene.
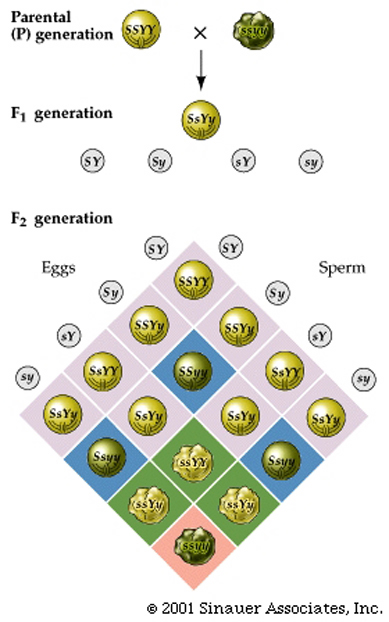
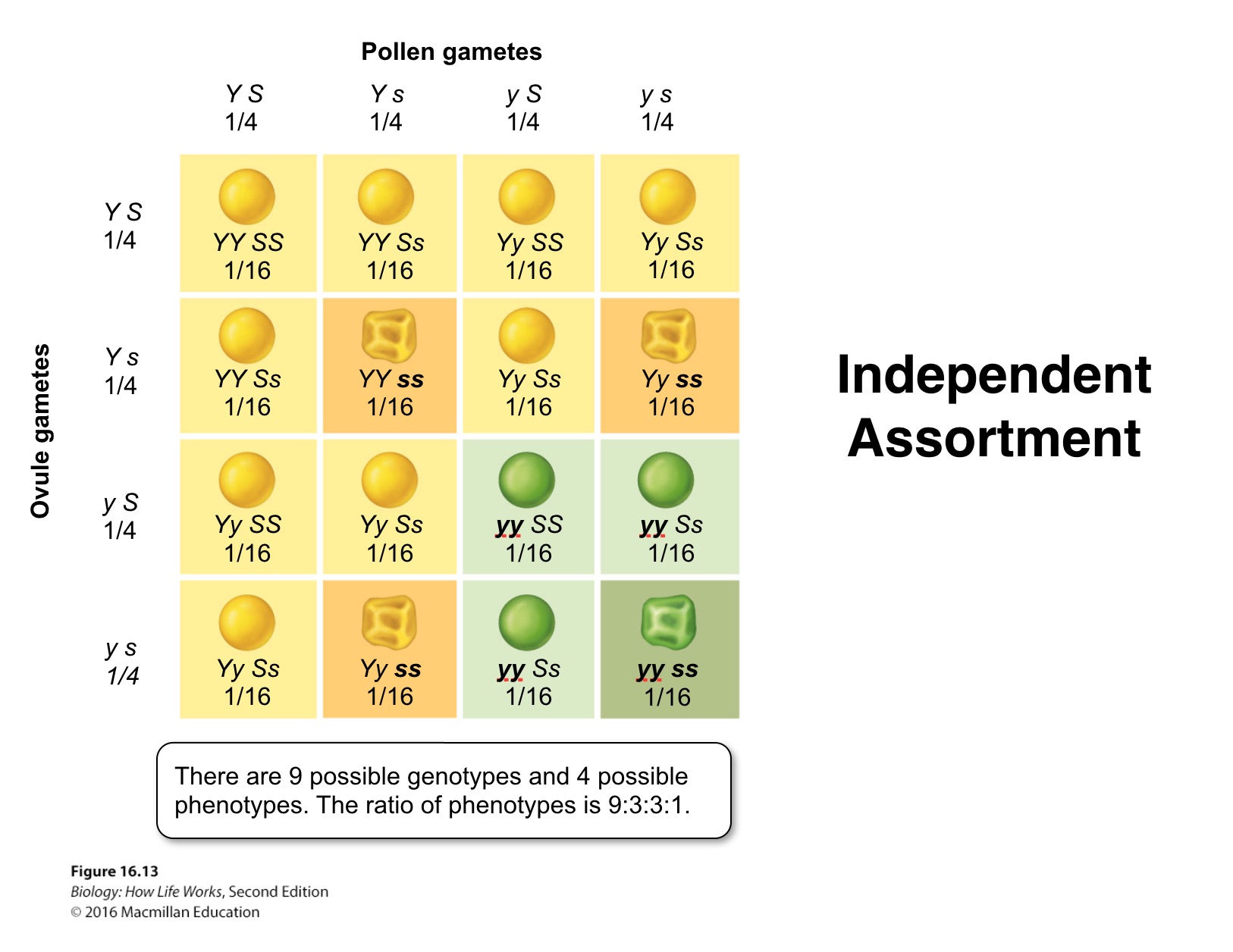
In this cross, Four different gametes are possible and will be produced in equal proportions: SY, Sy, sY, and s y.
Random fertilization of gametes yields the outcome visible in the more complex Punnett square.
{kind=link}
Note that it is now 4 x 4 table construction to accommodate 16 possible genotypes.
Filling in the table and adding the like cells reveals a 9:3:3:1 ratio of the four possible phenotypes (smooth yellow, smooth green, wrinkled yellow, and wrinkled green).
Monohybrid crosses, dihybrid crosses -and even more complex- trihybrid crosses: same principles.

You should know how to test for Mendel's laws.
Remember, Mendel had no knowledge of this or meiosis. Even so, we now know that meiosis nicely explains his law of segregation...
Mendel looked for characters that had well-defined, alternative traits and that were true-breeding.
While the animation of meiosis (above) provides a lovely mechanism for Mendelian inheritance that satisfies BOTH his laws, for this to apply to ALL of Mendel's "well chosen" traits they would all need to exist on different chromosomes, which (in hind-sight) we now know, is not the case.

How can his 2nd law of independence apply to "traits" that are carried on the SAME chromosome????
What about those "Chiamata"??? If the probability of a single chiasma forming between two different traits / genes is "ONE" (i.e. it is a certainty), then the assortment of alleles would, in essence, be the same as if they were on separate chromosomes (i.e. random).
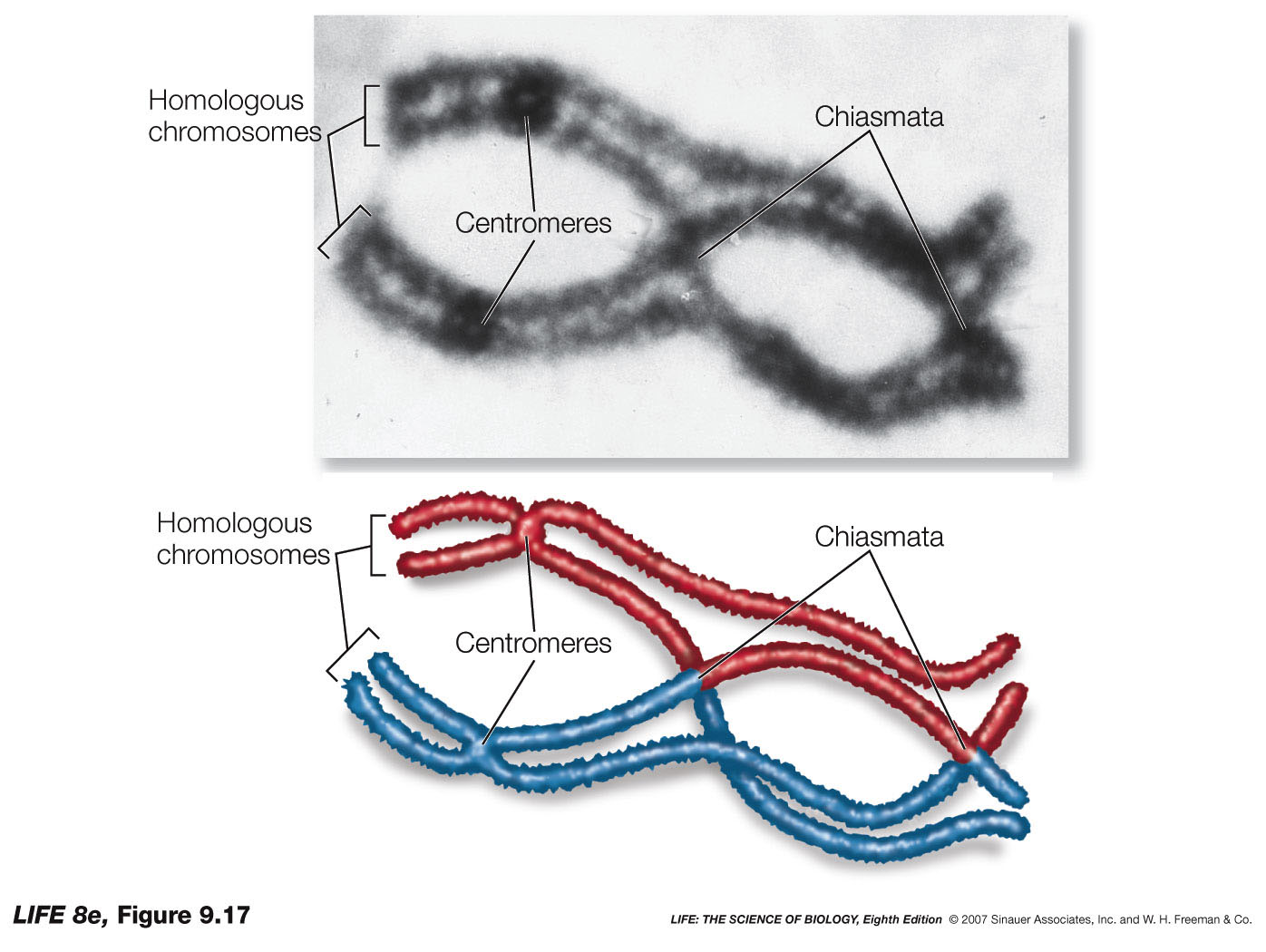
While Mendel could not comprehend any of the genetic intricacies that his laws might suggest, his determination of his abstract "unitary" traits had far reaching implications in understanding inheritance of these genetic traits and how they could be inherited.
As we have discussed, each of the "gene variants" that Mendel called "factors" are termed Alleles and ALL variants of a given gene reside at a specific locus on a given chromosome.
Variation in allelic types arise by mutation or change in the DNA sequence.
Allelic variation,
therefore, can arise due to a mutation
at a given locus, and (if phenotypicaly different from the original version can be analyzed through classical genetics -once that variation has been incorporated
Alleles can be randomly mutatagenized
to become a different allele, depending
on DNA sequence changes and then potentially maintained or lost -again in a random manner- unless
acted upon by a selective pressure.
"Wild Type", as we have discussed prevously, is a term used to demonstrate the most common alleles / traits that are present in a given population. Please be aware that, being the "most common" allele in a population does not always mean that the wild-type allele will be the dominant or "superior" allele.
However, some definition of "normalcy" in the genetic expression of traits does provide an incredible advantage in the understanding of how genes work in the whole organism...
One particular, rather grim, but striking example of this would be "Huntington's Disease", which is an inherited condition wherein nerve cells in the brain break down over time. It typically starts in a person's 30s or 40s. Usually, Huntington's disease results in progressive movement, thinking (cognitive), and psychiatric symptoms. No cure exists, but drugs, physical therapy, and talk therapy can help manage some symptoms. How and when the disease starts to effect the indiviudal is dependent on the presence of the allele. Presence of the trait as Hh (heterozygous) is the ususal form in which it begins to manifest itself in mid adul hood. However, if present in the homosozygous DOMINANT form, HH, the disease is more pronounced and manifest much earlier on -often in pre-adolescence.
In the early twentieth century, G. H. Shull crossed two varieties of corn, and the yield went from 20 to 80 bushels per acre, thus defining a, now common, agricultural practice to increase production in plants.
{kind=link}

This is called either hybrid vigor or heterosis, whereby crossbred strains show superior qualities, and it is a result of the fact that continued genetic "in-breeding" often leads to deleterious traits becoming "fixed" in the gene pool (an aspect we will address later). The mixing of two strains, therefore, reintroduces a number of selectible traits which can potentially enhance the gene pool of the newly defined strains... Use of such hybridization techniques has been a common agricultural practice ever since -to increase production in crops and broadened to cattle etc...
An hypothesis called overdominance even proposes that the heterozygous condition -in certain important genes- is often considered to be selectively "superior" or have a higher fitness than the presence of either homozygote. This occurs far more frequently in cases where a single trait controls more than one phenotype. Such heterosis, however, would be slightly at odds with Mendel's idea of "independent" traits.
Even so, given the potential of heterosis (cross bred alleles show some qualities that can be superior to their parental counterparts), this would threaten the very crux of Mendel's ideas which are premised on the "particulate" nature of genes, and where one allele in any given gene pair is simply dominant over the other
Perhaps, Mendel wasn't perfect...
No, he wasn't, he was also only observing, NOT deducing. Even so, the fact that we are still talking about him is testament to how much he did get right.
EXTENSIONS or "Modifications" to Mendelian Genetics -tend to vindicate Mendelian Genetics at the genotypic level.
Same basic scheme, but different phenotypical expression......
Heterozygotes may show an intermediate
phenotype.
For example, red-flowered snap dragons when crossed with white snap dragons will generate
pink-flowered plants.\

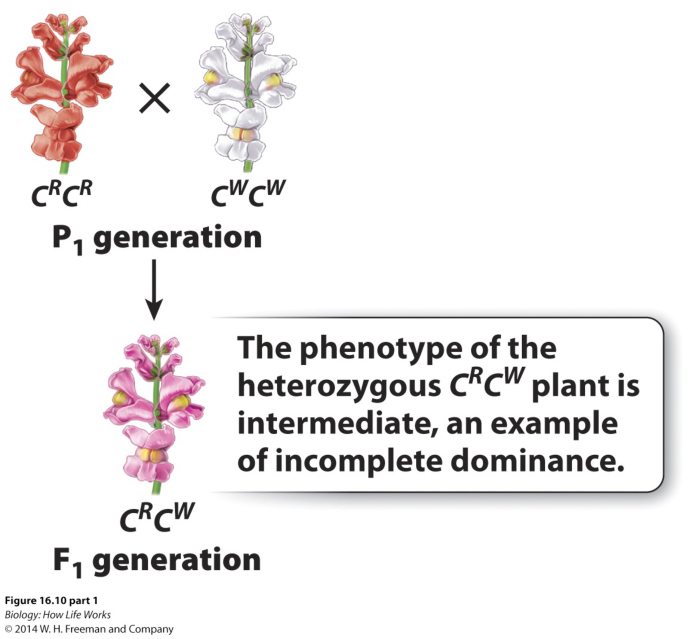
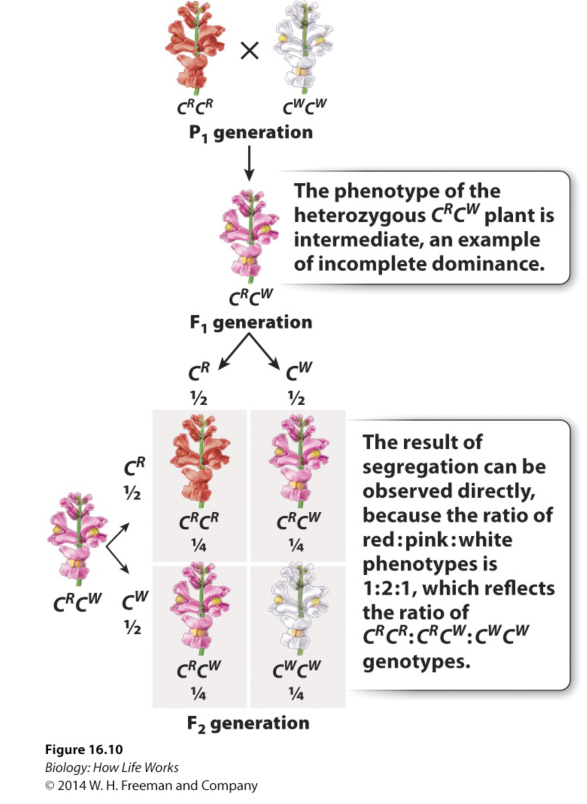
While this phenotype (on the surface) might tend to support the blending theory that Mendel was so avidly trying to overcome, the F2 progeny, of a "selfing" of the pink flowers admirably demonstrates, Mendelian "particulate" genetics, the reason for the change in colour results from a penomenon called "incomplete dominance".
Another example, where Mendelian Genetics apparantly falls down, can be seen in blood types. In this instance, however, Mendel’s laws are also not compromised -at the genetic level.

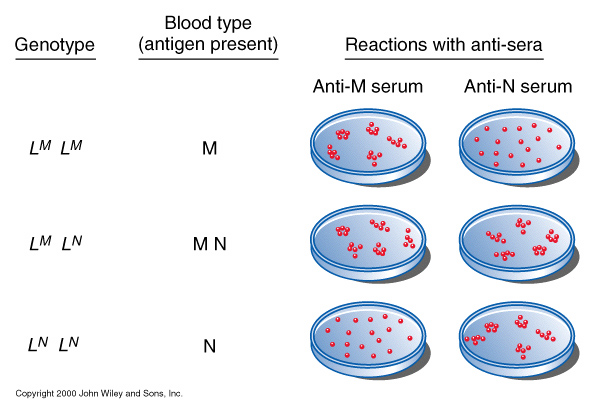
Cemelias, cows and even blood-types *see figures above) are said to exhibit codominance, where both alleles are expressed. Note that in codominance the phenotype of the heterozygote is completely different (not just a "blend" of the two homozygotic phenotypes).
Mendel made a series of artful choices in choosing traits in peas that "just happened" to be examples of complete dominance.
Consequently, Mendel analyzed his "units" in terms of dominant and recessive traits, there are many examples, where variations at a single locus, gives rise to multiple phenotypic effects.
So, let's look a little closer at the behoaviou of genes and their alleles.
Alleles
and their Interactions
Differences in alleles of genes
are often slight differences in
the DNA sequence at the
same locus, which result
in slightly different products,
and can give rise to different phenotypes.
Many "genes" can have "multiple alleles".
A population might have more than two alleles for a given gene, even though only a maximum of TWO can be present in any given "diploid individual".
In rabbits, coat color is determined by one gene with four different alleles. Five different colors result from the combinations of these alleles.
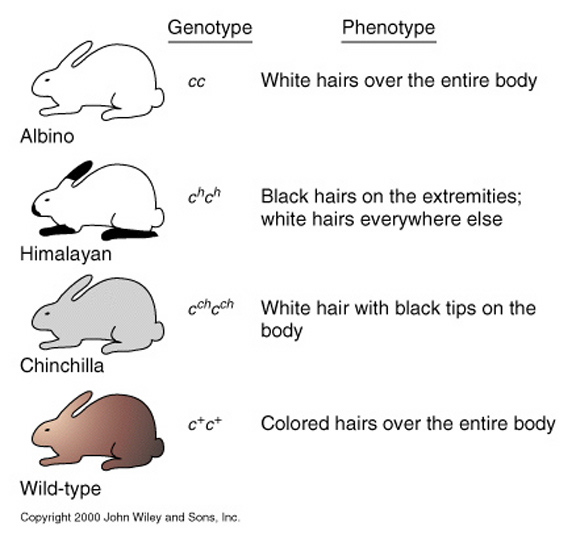
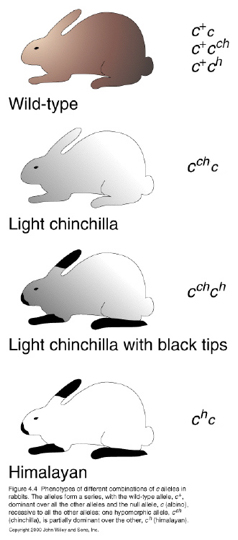
Again, as can be observed, Dominance need not be complete.
Multiple Alleles:The "ABO" blood-type system
The ABO system is now known to be a polymorphism of complex carbohydrate structures comprised of glycoproteins and glycolipids expressed at the surface of red blood cells.
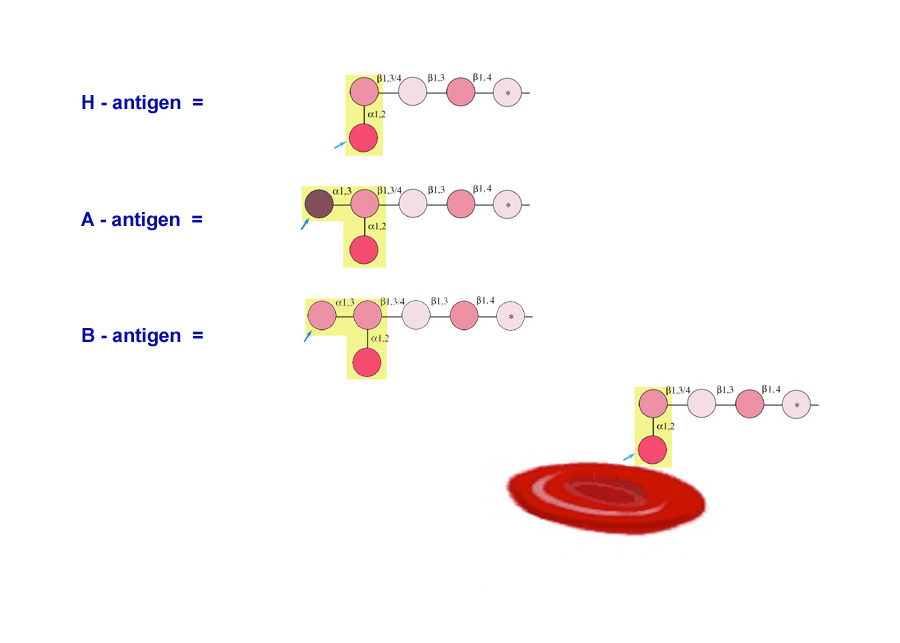
The alleles for blood type are IA, IBand IO. They all occupy one locus.
These alleles determine which antigens (proteins) are present on the surface
of red blood cells.
These "antigens" react with proteins called antibodies in the serum of certain individuals and cause the blood to clot.
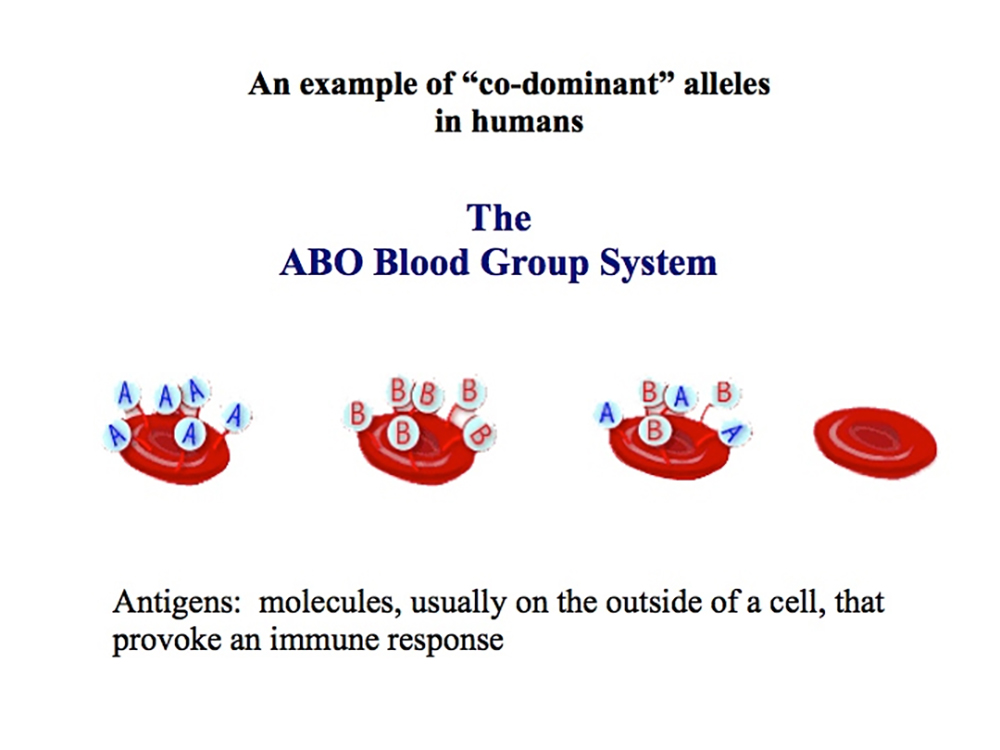

The "ABO" blood-type system, however, is not just a series of antigens on the surface of each of red blood cell, but also feature their corresponding "antibodies" that exist within the blood serum.
If A antigen (blood cell) and A antibody (serum) are present in the blood... the blood will clot!!.
Therefore, the result of mixing blood from each of the different ABO blood types can result in red blood cell agglutination, or clumping, which may prove to be fatal.
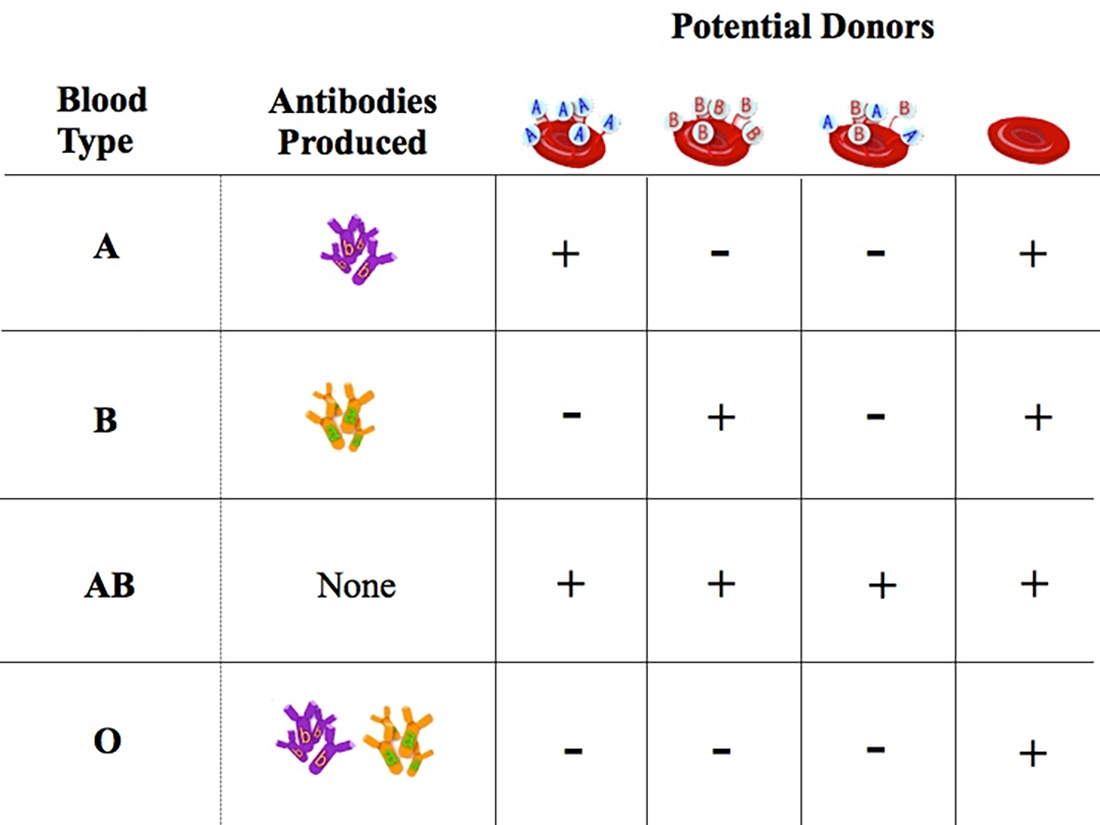
| Blood Type | Genotypes | ABO
Enzymes Present |
RBC
Antigens Present |
Serum Antibodies |
|---|---|---|---|---|
"A" |
IA IA,
IA IO |
"H", "A" | A, H | anti-B |
"B" |
IB IB,
IB IO |
"H", "B" | B, H | anti-A |
"AB" |
IA IO |
"H", "A", "B" | A, B, H | none |
"O" |
IO IO |
"H" | H | anti-A,
anti-B |
Chart Of A-B-O Blood Donor & Recipient Compatibility
|
Note that blood-types A and B can be considered to be "codominant", as the presence of both alleles AB gives rise to a completely different phenotype, when compared to a homozgous AA or a BB equivalent.
What about Rhesus factors?... these are a whole new class of antigens, discovered when blood from "rhesus monkeys" was injected into guinea pigas (circa 1940's). There are over 50 different types of Rh factors, but the most comomonly known one is the D antigen (Rho[D]), which -if it is present- indicates that that person is Rh-positive; if the D antigen is absent, that person is Rh-negative.
In contrast to the ABO system, however, antibodies to Rh antigens aren't necesssarily inherent to the person's blood, and can develop as an immune response after a blood transfusion, or during pregnancy.
At a mother's first antenatal screening, blood tests are taken in order to determine her blood type (A, B, AB or O) as well as her rhesus status (Rh-positive or Rh-negative).
If the mother has the rhesus factor (which is a protein on the surface of her red blood cells) then she is said to be Rh-positive. If not, then she is Rh-negative. 85% of people are Rh-positive.
The rhesus state only really begins to play a role during pregnancy if the mother is Rh-negative, the father is Rh-positive and the baby is also Rh-positive.
Rh(D) positive cells contain the D antigen, which can stimulate Rh(d) negative blood to produce harmful antibodies that can destroy red cells. The harmful antibody is called ‘anti-D’ antibody, and can be produced by a mother who is Rh-negative carrying a baby who is Rh-positive.
Rhesus incompatibility doesn’t cause any problems with a first pregnancy because (unlike the AB antibodies) the rhesus antibodies aren’t inherently present in the mother’s blood.
However, in subsequent pregnancies, if the babies are Rh-positive, there may be a problem. The mother’s antibodies can / will cross over the placenta into the baby’s blood causing a reaction.
This causes problems with the baby’s haemoglobin levels (the iron-carrying element in the red blood cells) which could then fall, causing anaemia. Blood transfusion would then be necessary (see chart below) at birth and the babies could also be severely jaundiced.
Gene Interactions
Some genes give rise to gene products
that alter the
effects (phenotypes) of other genes.
"the interaction of genes that are not alleles, in particular the suppression of the effect of one such gene by another."
Given that genes of any organism do not operate in total isolation from one another, but obviously are functioning in a common cellular environment. a phenomenon that occurs when the alleles of one gene covers up (or alters) the expression of alleles of another gene.


Labrador coat colour is an example of Epistasis in that there are two genes which affect coat colour -the "E" gene, which affects the presence of a dark pigment in the coat and the "B" gene, which governs the degree to which that pigment is present in the coat, where B gives rise to a "black coated lab", but only if the genotypes at the E locus are E_ or EE.
While blood groups would also be a more than adequate example of this phenomenon, a quite overt example of epistatic effect, albeit more indirect, would be life and death consequences!

In this instance the same gene that affects coat colour in the mouse also has some influence upon development of the embryo. Thus, one gene, does not always mean one function. If, as in this case, It has more than one function and is therefore considerred to be pleiotropic.
Some might even argue that this is not technically "true" epistasis... Why? what do you think?
Ratios are important characteristics: 3:1 (and 1:1 -test cross) for monohybrid crosses and 9:3:3:1 for dihybrid crosses.
Extension of Mendelian Genetics:-
Another example is fur coat colour in mice can be seen in the example shown in the picture:
The B allele determines a banded pattern, called agouti.
The recessive b allele results in unbanded hairs.
The genotypes BB or Bb are, therefore, agouti.The genotype bb is coloured solidly black .
Another gene, at an entirely different locus, determines if any coloration occurs at all.
The genotypes AA and Aa have "Agouti", whereas the double recessive aa are "solid coloured.
Mice that are heterozygous for both genes are agouti.
An F2 phenotypic ratio of an initial parental cross between a BB, AA and a bb, aa would be: 9 agouti: 3 black: 3 cinammon and 1 brown .The corresponding genotypes are 9 agouti (1 BBAA + 2 BbAA + 4 BbAa):3 Cinamon (1 bbAA + 2 bbAa): 3 Black (1 BBaa + 2 Bbaa + 1 brown (1 bbaa)
A 3rd locus "C", controls whether or not there is "any colour or not, and if present in the recessive form (cc) then this grouping will not allow ANY colour to show through -agouti, brown, cinammon or black- the miice are considered to be "albino".
Other examples of gene/allelic expression, which differ from expected patterns of Medelian inheritance are:
Duplicate genes, occur when two distinct genes affect the same phenotype in the same way.
One of the best examples of this would be the shape of the shepherd's purse pollen sac.
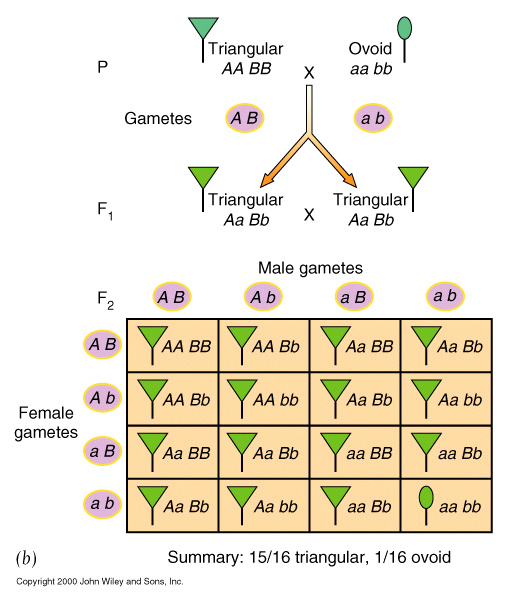
Complementary gene activity, occurs when two distinct genes affect the same phenotype in a complementary way, whereby the presence of BOTH dominant alleles are required to give a particular phenotype that is distinct from either being present by itself.
Polygenes also mediate quantitative inheritance.
Individual heritable characters
are often found to be controlled
by groups of several genes, called
polygenes.
Each
allele intensifies or diminishes the
phenotype.
Variation is continuous or quantitative
(“summation" or "adding
up” of all the traits).
Examples of continuous characters
are height, skin color, and (possibly)
intelligence.

| Incomplete dominance: | eg. Four o' clocks, carnations. BLENDING 1 : 2 : 1 |
| Codominance: | eg. M and N
blood groups on
chromosome 4 having
specific antigens
M and N
1 : 2 : 1 but where the heterozygotes (MN) gives rise to a distinct phenotype |
| Multiple alleles | ABO blood types, A and B are dominant to O , but A and B codominant to each other |
| Epistasis | essentially "eliminates"
or "masks" phenotypic
expression of other
genes,
eg. Labrador dogs fur colour, albinoism in mice |
| Lethal alleles | eg. Yellow colouration in mice fur. 2 : 1 . Pleiotropy(?) |
| Several genes / same character | Coat colour in mammals,
eg. mice A (agouti), B (black/brown), C, (colour) D (intensity), S. (distribution) genes |
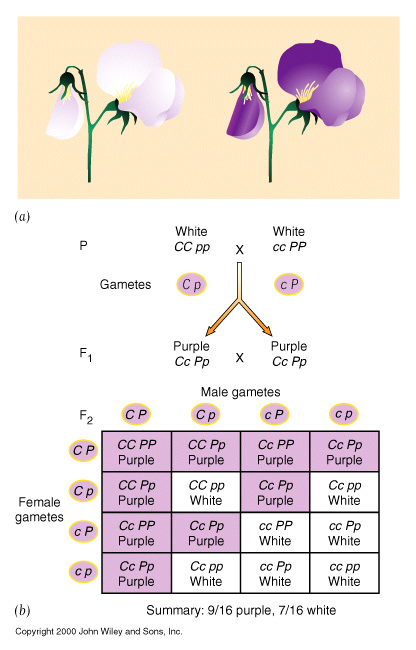
| Complementary gene activity | eg. Pea plants, purple colouration. 9 : 7 |
| Duplicate gene activity | eg. Shepherd's
purse, Round over
narrow fruits, where
both A 1 - and A 2 - can cause heart shape 15 : 1 |
Nature
vs. Nurture?
When thinking about the importance
of either genotypes or phenotypes,
one must always consider the immense
importance of.....the environment , and how it can affect
the action(s) of any given gene.
Variables such as light, temperature, and nutrition can dramatically affect the translation of a genotype into a phenotype.
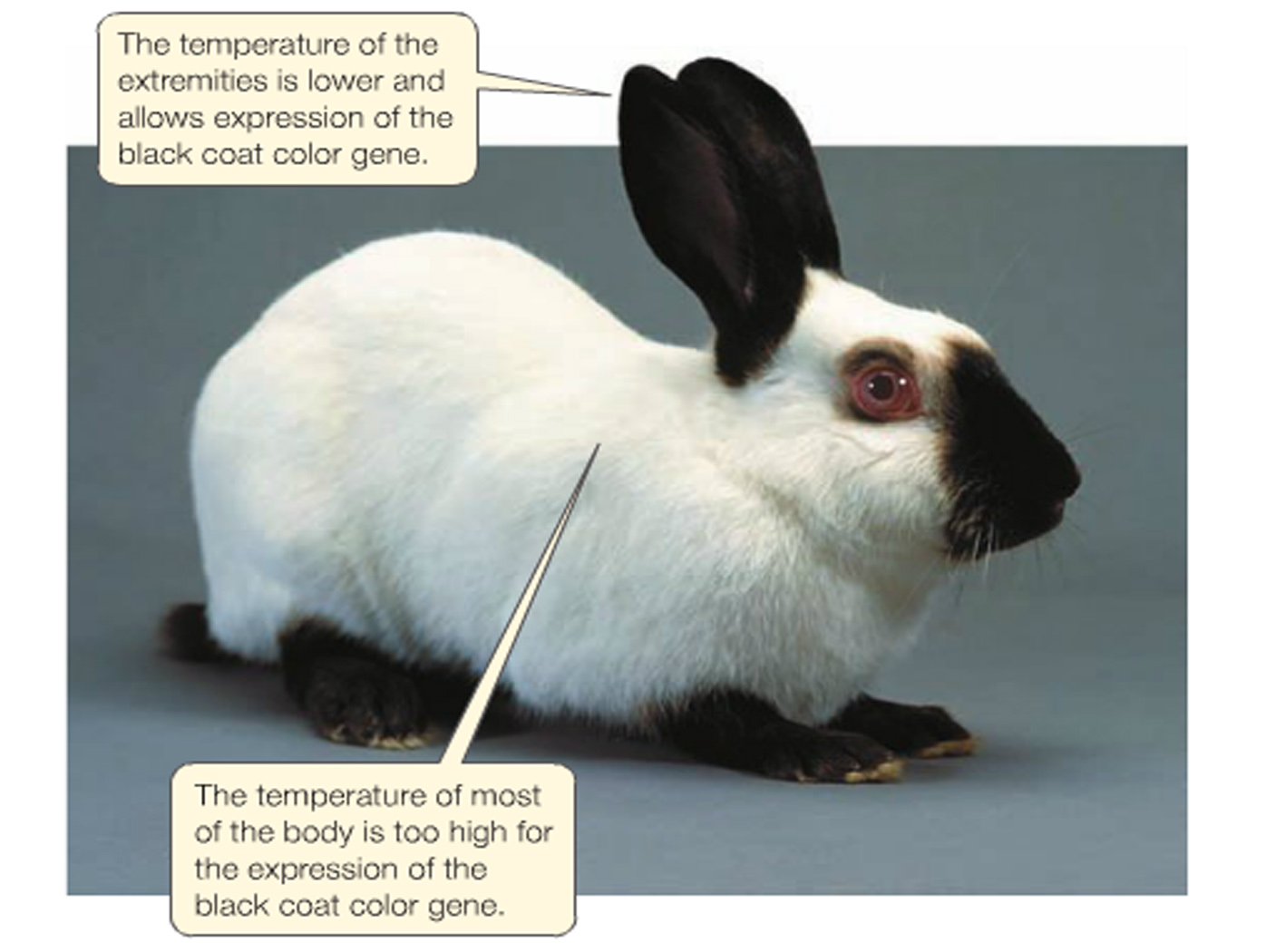
For example, the darkness of the fur on extremities of a Siamese cat is affected by the temperature of that region. Darkened extremities normally have a lower temperature than the rest of the body.
Such colouration can be manipulated experimentally.
Gene linkage and the role of the sex chromosome:
Mendel did all his analyses with plants, which -like many other plants- exhibit BOTH male and female reproductive structures in the same adult plant (termed monoecious, or "all in one house").
{kind=link}
What about animals and plants that have individuals which are either one or the other sex (termed dioecious, or having “two houses”).
In most dioecious organisms, the sex of the organism is determined by differences in the presence/absence of a set of chromosomes, or the presence/absence of discrete chromosomes.
In honeybees, eggs are either fertilized -and become diploid females- or they are not fertilized -and become haploid males, drones.
In Mammals, such as humans...
Humans have different sex chromosomes, X and Y. Males have X and Y; females have X and X.
Again, the sex of the offspring is also determined by the sperm.

About Contact GSU Academics Degrees & Majors |
Admissions Undergraduate Research Dept. of Biology |
Libraries University Library Campus Life Housing |
Athletics Alumni
|