

- Home
Welcome !
Welcome to john houghton's home page for his biology courses. This site is designed as a hub for curating and sharing lectures, course syllabi, assignments, and links to relevant resources. Use the menu bar at the top of the screen to navigate through the site.
(Please note: this page is currently under construction.)

- BIOL 2107
Fall '23 CRN86772
Lectures: (1)
- Courses
BIOL 2107 Principles of Biology I

| Lecture 4: Origins and Mechanisms of Evolution ctnd. |
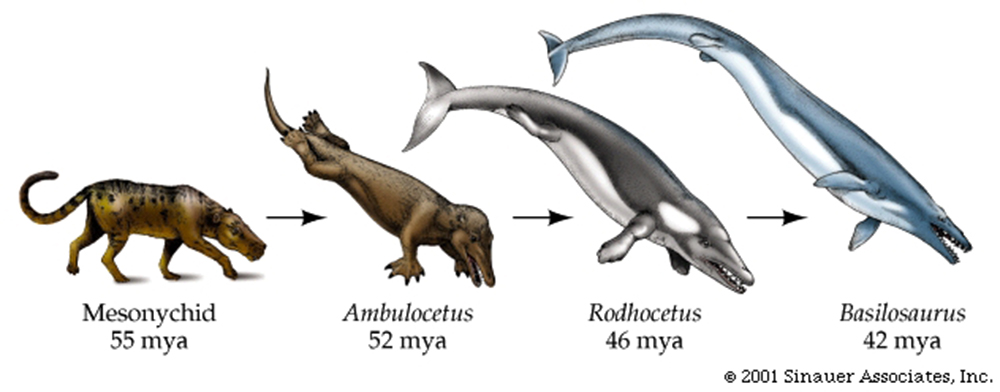
Evolution of Whales ? -MOVIE Lucy ? -MOVIE
Selection Types -MOVIE Genetic Drift -MOVIE D K P C O F G S -MOVIE
Fossils records show that many evolutionary changes are also gradual (in geological time). While these changes do explain some of the changes that have occurred through geological time, they also demonstrate that an incomplete record -in a similar way to faulty reasoning (such as circular arguments or the use of "invalid probes") can sometimes falsely suggest or even conceal times of rapid change, or completely redirect potential heritable linkages.

But we are getting a little ahead of ourselves....
In the early to mid 19th century, at about the same time that a researcher called Sir Charles Lyell was appreciating the idea that the age of fossils somehow related to the age of the rocks in which they were found......
Darwin began his "Voyage of the Beagle" (1831)
and his first opus: The Origin of Species 1859, and the crucial determination.....
As a result of his travels, Darwin developed his idea of "evolution by natural selection" by carefully observing nature, especially during his voyage around the world, and comimg up with a rationale for how he understood that these variations could exist..
He based his theory (little "t") on well-known facts and some key inferences. Obviously, he had no examples of the action of natural selection, so he based his arguments on his current knowledge of artificial selection by plant and animal breeders of the day.
Modern Genetics has since elucidated the mechanisms of heredity, which have provided a solid base that supports and substantiates (to a large extent) Darwin’s theory. These findings, along with others over the subsequent years, began to change the "t" into a "T".
In essence, Darwin "observed" various traits of the wild life around him, both in South America and on the islands just off the coast. He noted, similarities and differences, and wrestled with the ideas of how these similarities AND differences came about. He understood, the importance of potentially "informative", shared traits, and that the differential loss and gain of these itraits could lead toward distinctions and similarities.
Moreover, HOW these traits change can also be highly informative.
Natural selection, which is the "selective" process by which variation of heritable traits can determine the success of the "reproductive efforts" of animals (life foms) within a given population, and lead to changes in the genetic makeup of indivdiual organisms within that population.
All similarities and differences within and among different types of animals, therefore, when taken together, can provide a series of "near" and "distant" relationships... -ultimately forming family trees, which we will discuss at a later date.
Some of the traits that are shared among animals and plants of various species can be considered to be "informative" in that they can be used as exceptionally reliable evidence that allows evolutionary scientists to deduce (infer!!) heritable relationships and potentially build ancestries for animals that may otherwise not be fully understood.
(MOVIE) Whale evolution.....from wolves or "wolf-like creatures"?
and (MOVIE) Lucy .......
Three important thoughts that I would like to stress from having reviewed the movies.
a) Evolutionary change in whales is not like the Tarzan movie at the end of the 90's.....
b) The use of "shared ancestries".... It is important that we understand that, according to the fossil records whales have probably descended from a "wolf-like" animal NOT from a "wolf". Likewise "Lucy" is not an "ape" but an "ape-like hominid".
b) That the inference of bone configurations of modern (latter-day) analogues -allowing for adaptation over time- really can be used to relate the evolutionary development of known organisms.
Ideally all of these ideas of change(s) reflect back upon Darwin's appreciation of the inhabitants of the Galapagos islands, and the finding that there are forces at work which affect "if" and "how" different animals/plants in a given population are able to survive.
Of course all this evidence pre-supposes "honesty" among scientists......which is not always the case, either as a consequence of being unable to interpret the data correctly, or even as a consequence of outright fraud.
What is also important to note, however, is that even a superficial understanding of these concepts allows us to appreciate that the primary forces of evolutionary change exert themselves, not on individual organisms, but on populations of related individuals.
Also implicit within this, and any understanding of evolution is that this "population of individuals" is not a number of identical clones, but a group of individuals that exhibit similar phenotypic traits (the operative word being "similar"), and that each individual has a slightly different organismal "signature" to donate to the collective "gene pool".
To put Darwin's ideas in a 21st century context with a little more clarity (gained from almost 190 years of hind-site), let's define some of the vocabulary that we might use to more readily understand some of the ramifications (consequences) of Darwin's ideas.
- population: all individuals of the same species occupying the same area.
- gene: a unit of heritable information -usually associated (at the molecular level) with a specific region located on the chromosome.
- allele: - one of two or more slightly different forms, or "variants" of a given gene.
- genotype: a selection of the genes that make up an individual.
- phenotype: the consequence(s) of all the allelic interactions that give rise to a visibly determinable "type".
- gene pool: all the genotypes within a population.
- allelic frequency: the frequency in which an allele shows itself in a given population or gene pool.
From and understanding of a few of these fundamental concepts we can then build a greater undrstanding of more complex evolutionary terms, such as-
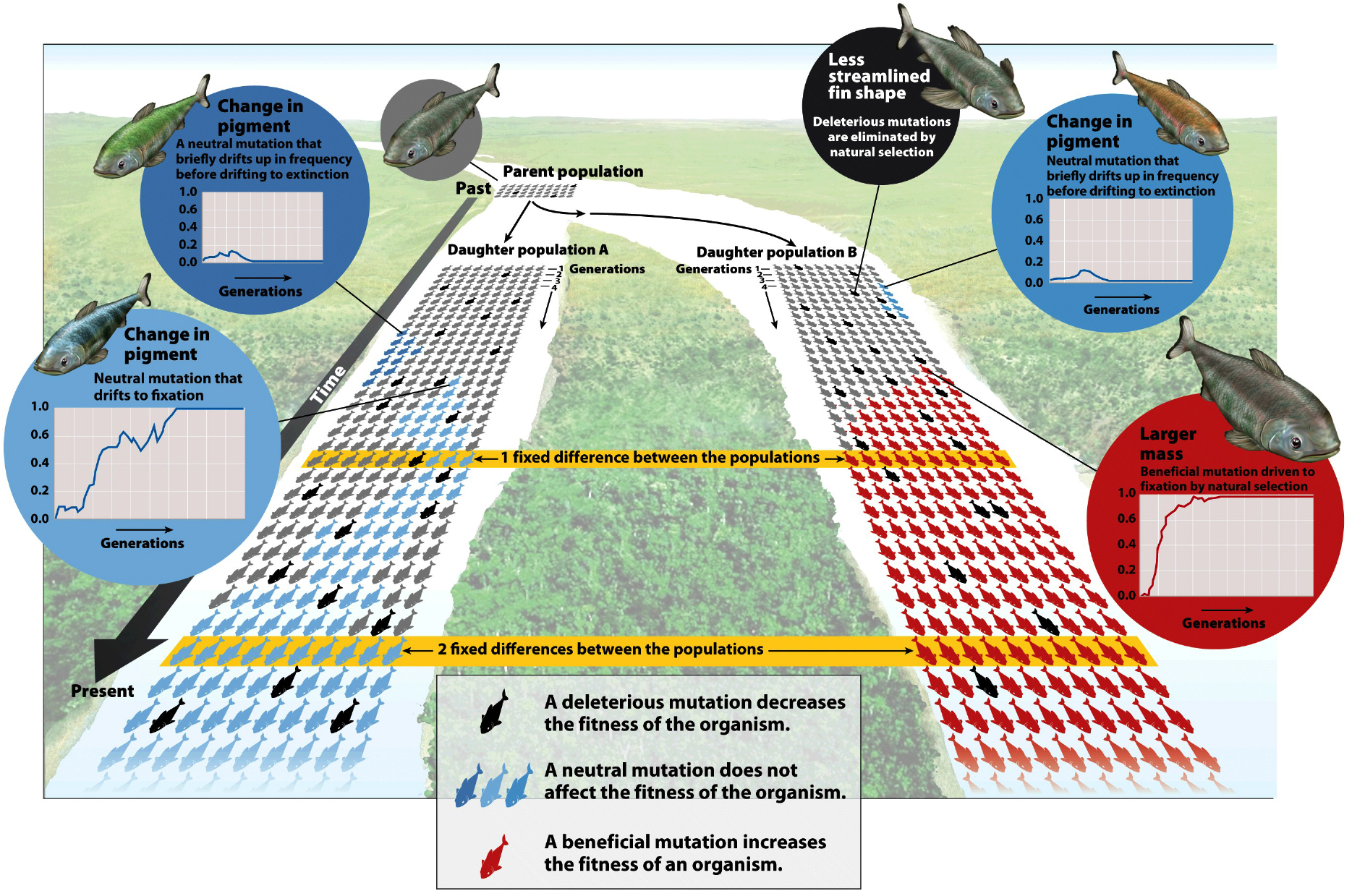
Genetic drift: a random change in allelic frequency over time and appreciate this as being a key mechanism of evolutionary change.
As a result, the effects of genetic drift might be strongest in small populations (?)
Why would that be (?) -the fewer individuals in the population, the more likely it is that random fluctuations will completely disrupt the allelic frequency.
In the short term (i.e over a few generations), one might expect allelic frequencies to increase and decrease in a random, unpredictable way, as a result of genetic drift .
Consider a newly introduced allele within a population. The presence of this allele will have an allelic frequency that can change over time. At some stage, perhaps, the frequency of a particular allelewould become fixed in a population or lost from a population for one reason or another, but this need not happen -especially in large populations.
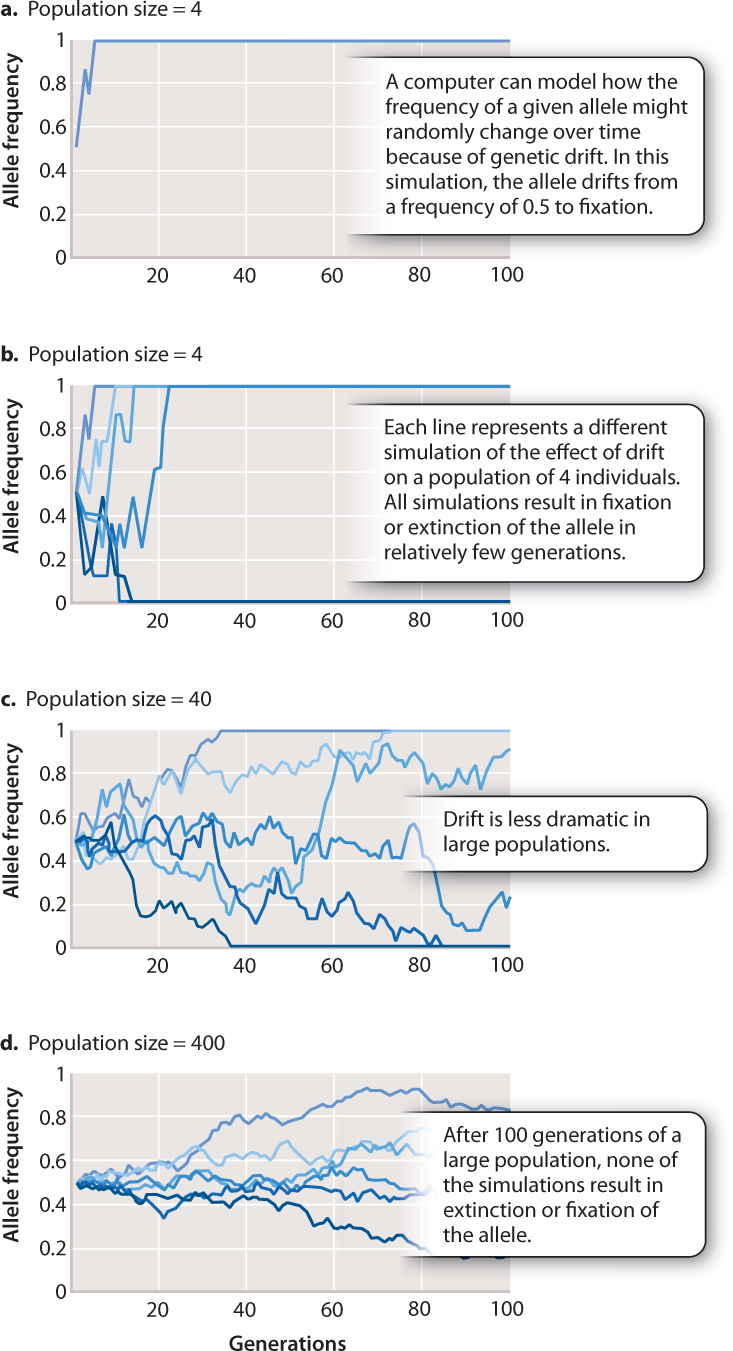
If these traits are selected for (or against) the rate at which these allelic traits become more fixed (or lost) within a population is increased. Again, changes in the number of individuals within a population that carry any particular allelefor a given trait is more likely to be affected in a small population than it is in a large population.
In the longer term, the major consequences of genetic drift lead to a homogenization of phenotypes within a population (everyone starts to look alike), especailly if there is some external selective pressure as certain alleles (read "variations") will be lost as others become "fixed". Thus, genetic drift gives rise to a loss of variation in the gene pool (genetic variation).
Now couple genetic drift with Darwin' s idea of "natural selection".... and you can begin to see how the effects of inherent populational variation and fluctuatios in that variation (along with environmental conditions) can begin to alter the allelic frequency within different populations over time....
 ;
;  .
.
As a counteraction to all these chages is the notion of - Gene flow(?) which would tend to INCREASE genetic variation within a given population.
Gene flow is the intermingling of separate traits among similar populations. This increase occurs because individuals from other populations will bring in alleles that would otherwise be absent or rare (may be even lost) from the population that is being observed. In other words they would add variety to the gene pool.
Note that the effect of gene flow on genetic variation within a population is the opposite of the effect of genetic drift.
In the short term (i.e over a few generations), one might expect allelic frequenciesto increase and decrease in a random, unpredictable way, as a result of genetic drift
As we ave just discussed, however, in the longer term, the major consequences of genetic drift lead to a homogenization of phenotypes with in a population.
On the other hand, gene flow(?) would tend to increase genetic variation within a given population.
Quick question. How might you think honey bees contribute to the gene flow among assorted pollenating flowers?
In coining the word and idea of "natural selection" Darwin was able to provide a potential mechanism by which different phenotypic traits changed (and how they continue to change), an understanding of which, can be highly informative.
In the modern terms
Natural Selection can be understood to be:
"the gradual process by which heritable biological traits become either more or less common in a population as a function of the effect of inherited traits on the differential reproductive success of organisms interacting with their environment".
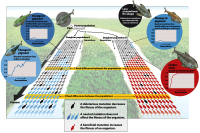
There are also classic examples of other more severe "external" factors that can have significant effects upon the makeup of any gene pool.

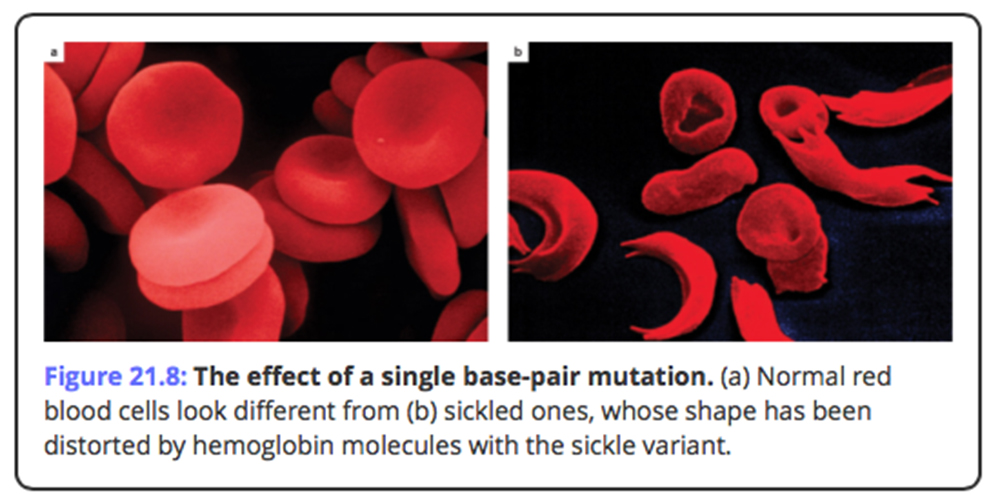
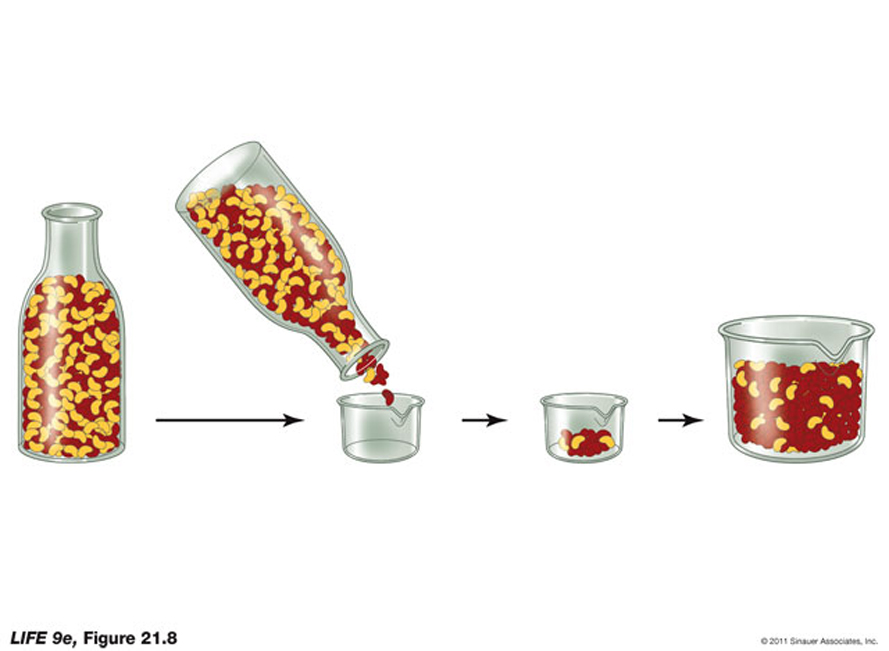
- bottleneck: severe reduction in population size due to intense selective pressure or a natural calamity or catastrophe, which alters the allele frequency...
As I was talking about in the last lecture.... over the life time of the earth there have been a number of "significant" events, such as meteorite collisions and indirect interactions with comets , which have had a cataclismic affect / change on climatic conditions -changing the potential for life to exist on the earth.
There is a considerable body of evidence to suggest that a meteorite may have been at least partially the cause of the somewhat abrupt mass extinction at the end of the Cretaceous period.. or alternatively...Volcanoes in the Deccan Traps
Are there Human bottlenecks??
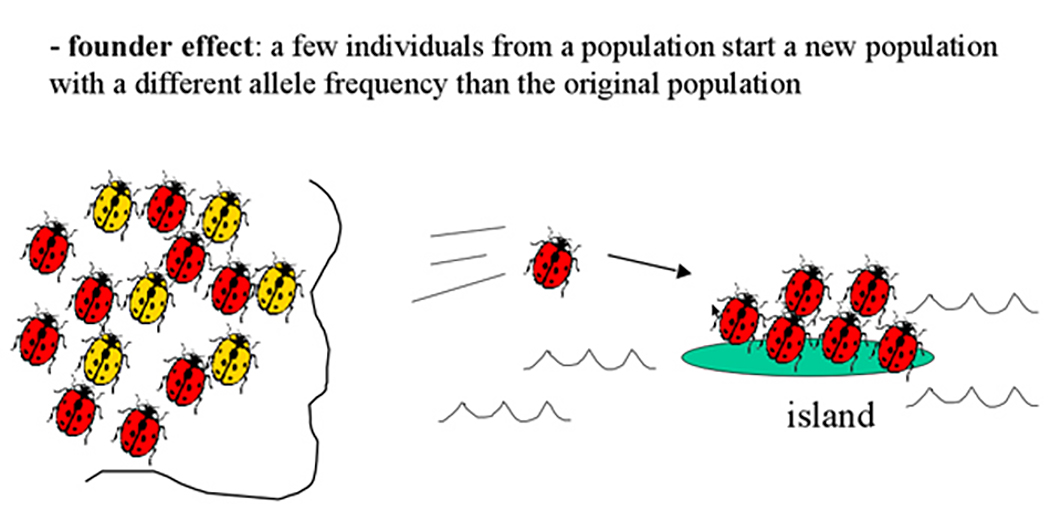
- founder effect: whereby a couple or even a single organism becomes "dislodged" from a population, and all genetic variation is limited to the individual or isolated individuals.
Human founder effects ... The Afrikaner population in South Africa and Huntington's disease traced back to 1652
Now, armed with all this "new" information (and vocabulary(?)), let's return to Darwin and some of his rather absurd notions that he wrote about in in his first book The Origin of Species, and let's ask a few questions .........
{kind=link}
QUESTION- Given what we have determined, might the "forces of evolution" work equally well on populations that have a limited gene pool, as they do in those populations with a much larger -effectively infinite variety of alleles within a its gene pool?
YES or NO... ?
This would explain the potential changes in the frequency of phenotypic changes (due to genetic drift) in animals / finches / tortoises that Darwin noticed on or in the Galapagos islands vs. similar changes (or a lack thereof) in their main land relatives.
Remember, Darwin sailed around the world, saw a great deal of flora and fauna, but it was the Galapagos islands that provide the important location for his idea of natural selection.
Really, all Darwin observed were phenotypic "variations of different shared phenotypes"
Question: Are there any constraints to evolutionary changes?
Answer: Yes.
At this stage, however, the scientific method and our understanding of these "constraints" upon evolutionary change can get bogged down in details.......
-------
Remember, implicit within this understanding of Evolution is that Evolution works on populations of "individuals" -each contributing a slightly different genotypic "signature" to the population's gene pool.
How did we get such a diverse gene pool?
Unfortunately, that question we will have to leave for another day -once we have dealt with Genetics and Mendal and his findings.
For now, I would like to ask how do we now adapt Darwin's evolutionary principles and begin to determine that there are sufficient differences between populations with "similar, but different, genetic makeup", and assign to each a different name,.....
....In essence how do we determine and then justify the term SPECIES that Darwin uses in his book, The Origin of Species, ?
Darwin and his contempories used appearance(s) (phenotype[s]) of animals/plants to define differences......often to distinguish one species from another, or (at the very least) recognize members of the same species present in different geographical locations.



Over almost 300 years ago, Carolus Linnaeus -following on from John Ray (> 350 yrs ago)- created the system of naming organisms that we still use today. even so, neither of these men were the first to do so, officially (remember Aristotle).
{kind=link}
Linnaeus classified over 4,000 organisms based on their physical characteristics and appearance, using a morphological concept of species, and his best-known accomplishment is creating a hierarchical system of organization for living organisms, using, what is called a binomial nomenclature. K P C O F G S
How many kingdoms are there....? (A) 3, (B) 4, (C) 5, (D) 6, (E) 8
So, Not K P C O F G S, but really - " D " K P C O F G S ???
Before this classification system was effectively put in to use, scientists had many names for the same organism, which -not too surprisingly- caused considerable confusion within the scientific community.
Even so, despite his revolutionary ideas (remember, Linnaeus' ideas preceded Darwin's reflections, which only began in the late 1800's ) scientists still clung to the belief that all species were present in their day, as they always had been.
After Darwin, howeve, this classification scheme needed significant refinement on precisely what was meant by each of the terms, (especially the term "Species" ) and how they came about.More modern, updates of Linnaean classifications arose in 1940, when Ernst Mayr proposed the "biological" definition of species, or the Biological Species Concept [BSC]:
"Species are groups of interbreeding (or potentially interbreeding) natural populations which are reproductively isolated from other such groups."
Problems....
One cannot apply the BSC definition to a considerable number of asexual populations.
Application of the BSC toward self-pollenating plants is (by its own definition) somewhat limiting.
Moreover, it is often very difficult to apply (practically) anyway. It requires breeding tests to verify the separation, but this is rarely used. Indeed, were breeding tests to be used to define such criterea, natural environments would potentially play an overiding role.
e.g. Do Homo erectus, Homo neanderthalensis and Homo sapiens represent the same or different species? This question is unresolvable using the BSC definition, especially when (as we now suspect) Neanderthals and Homo sapiens may have interbread.
Other Definition of Species include: The Phenetic (or Morphological) Species Concept -Cronquist (1988)- who proposed an alternative to the BSC that he called a "renewed practical species definition", where he very clearly defines species as
"... the smallest groups that are consistently and persistently distinct and distinguishable by ordinary means."
As you can see, determining if two populations are actually different "species"a can be quite a difficult task, especially as the very process of speciation is often a gradual process.
In a more current definition of the term "species", the concept of genetic integration has been coined, and used to differentiate between SOME types of species.
Genetic integration works on the premise that if individuals within a population mate with one another, but not with individuals of other populations, they are considered to be an "independent evolutionary unit", and can safely be called a Species.
But just how do/did New Species Arise?
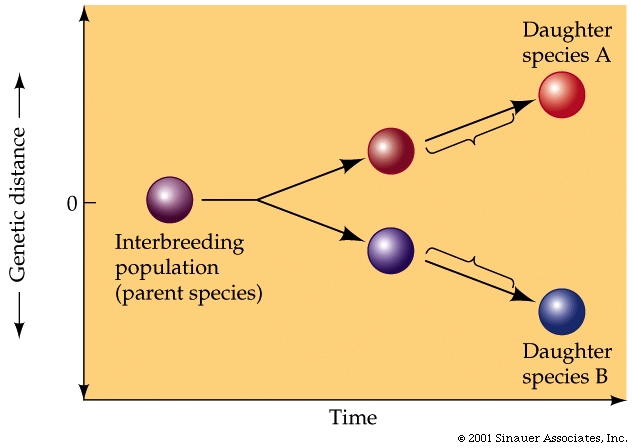
Speciation is the process by which one "species" splits into two.
We have already observed some possible mechanism of a potential speciation....have we not?...in the "black-bellied seed cracking finches of West Africa. But not all such evolutionary changes result in new species. The critical process in the formation of new species is the segregation within the gene pool of the ancestral species into two separate gene pools. Consequently, such speciation is often facilitated by an interruption of gene flow between or among different populations.
Allopatric speciation requires total genetic "reproductive" isolation.....or, when two or more parts of a single population become divided by a geographic barrier, alternatively known as geographic speciation.
Allopatric Speciation is thought to be the dominant form of speciation among most groups of organisms.
Even so, Darwin's seed-eating and bud-eating finches of the Galapagos archipelago demonstrate the importance of geographic isolation as a distinction that (along with population size, perhaps) can enhance the process of speciation.
However, the effectiveness of any barrier -geographical or otherwise- in preventing gene flow depends on the size and mobility of the species in question.
eg. an impenetrable physical barrier for a terrestrial snail may be not be as remarkable a hurdle for a butterfly.
Indirect evidence that most speciation among animals is allopatric is provided by patterns of species distribution.
These findings can lead to a tantalizingly plausible (but ultimately false) "aboslute" inferences......... if that geographic barriers always give rise to speciation....all organisms that are separated by a geographical boundary become species......long-distance romances don't work?
Unfortunately, it is not true...long-distance relationships can work.
Geographic isolation does not necessarily lead to reproductive incompatibility, as evidenced by the European and American sycamores.
Apparently, 20 million years of separation isn't such a long time if neither partner has to change too much.
Consequently, "physical" barriers need not be the only part of the "species" equation.
Anything that begins to separate the gene pool of the ancestral species will suffice.
Speciation, in the absence of any geographical barrier -or the subdivision of a gene pool when members of the daughter species are not geographically separated, but the two species become distinct as a function of inherent genetic polymorphism- is called...
Sympatric speciation.
Any example of a FUTURE(?) sympatric speciation springs to mind, perhaps??
(A) American Sycamores?
(B) West African Finches?
(C) Long necked Giant Toitoise in the Galapagos Islands?
(D) Birds and Bats?
(E) Red winged Blackbirds?
Additionally:
Fruit flies have speciated sympatrically in New York State for more than a hundred years.These fruit flies originally courted, mated, and deposited eggs only on hawthorn fruits.
About 150 years ago, large commercial apple orchards were planted in New York.
Some fruit flies began to lay their eggs on the apple trees, perhaps by mistake.
Consequently, their offspring sought out apple trees as adults and, therefore, mated with other fruit flies of similar heritage.
Today there are two sympatric species of picture-winged fruit flies in New York.

About Contact GSU Academics Degrees & Majors |
Admissions Undergraduate Research Dept. of Biology |
Libraries University Library Campus Life Housing |
Athletics Alumni
|