

- Home
Welcome !
Welcome to john houghton's home page for his biology courses. This site is designed as a hub for curating and sharing lectures, course syllabi, assignments, and links to relevant resources. Use the menu bar at the top of the screen to navigate through the site.
(Please note: this page is currently under construction.)

- BIOL 2107
Fall '23 CRN86772
Lectures: (1)
- Courses
BIOL 2107 Principles of Biology I

| Lecture 5: Speciation |
Implicit within our understanding of Evolution from the last lecture is that Evolution works on populations of "individuals" -each contributing a slightly different genotypic "signature" to the population's gene pool.
How did we get such a diverse gene pool?
Unfortunately, that question we will have to leave for another day -once we have dealt with Genetics and Mendal and his findings.
For now, I would like to ask how do we now adapt Darwin's evolutionary principles and begin to determine that there are sufficient differences between populations with "similar, but different, genetic makeup", and assign to each a different name,.....
....In essence how do we determine and then justify the term SPECIES that Darwin uses in his book, The Origin of Species, ?
Darwin and his contempories used appearance(s) (phenotype[s]) of animals/plants to define differences......often to distinguish one species from another, or (at the very least) recognize members of the same species present in different geographical locations.


Over almost 300 years ago, Carolus Linnaeus -following on from John Ray (> 350 yrs ago)- created the system of naming organisms that we still use today. even so, neither of these men were the first to do so, officially (remember Aristotle).
Linnaeus classified over 4,000 organisms based on their physical characteristics and appearance, using a morphological concept of species, and his best-known accomplishment is creating a hierarchical system of organization for living organisms, using, what is called a binomial nomenclature. K P C O F G S
How many kingdoms are there....? (A) 3, (B) 4, (C) 5, (D) 6, (E) 8
Monera / Archaea / Protista / Fungi / Plantae / Animalia / Insecta / Panera / Pianera -spot the wrong one(s)???
So, not just K P C O F G S, but really - " D " K P C O F G S ???
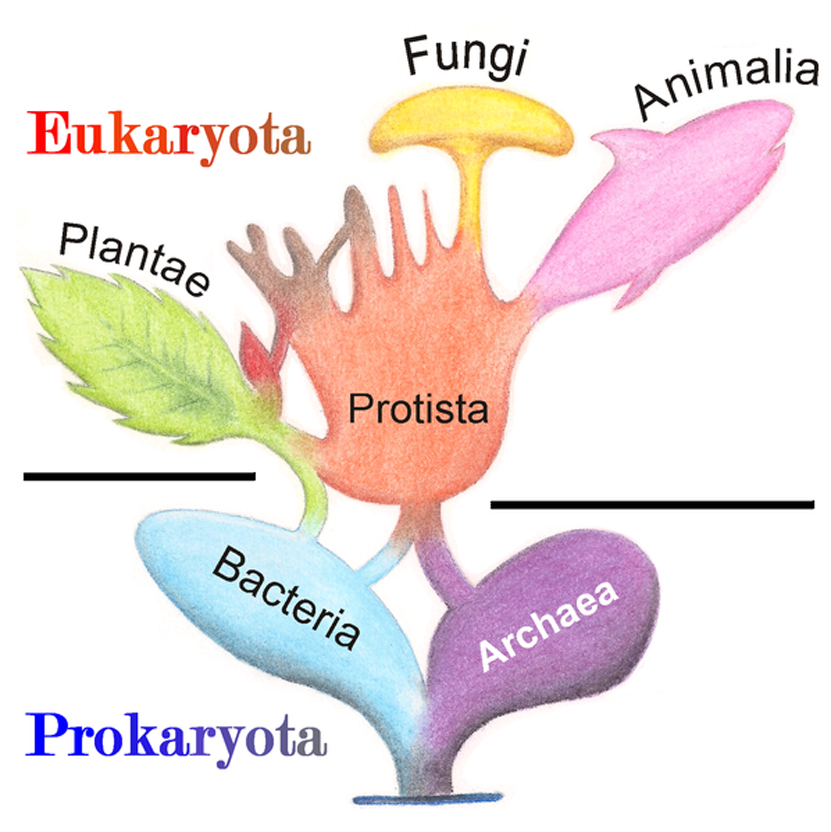
Before this classification system was effectively put in to use, chaos ensued, scientists worked in a "decentralized " void, using many names for the same organism, which -not too surprisingly- caused considerable confusion within the scientific community.
Even so, despite his revolutionary ideas (remember, Linnaeus' ideas preceded Darwin's reflections (which only began in the late 1800's ) scientists still clung to the belief that all species were present in their day, as they always had been.
After Darwin, howeve, this classification scheme needed significant refinement on precisely what was meant by each of the terms, (especially the term "Species" ) and how they came about.More modern, updates of Linnaean classifications arose in 1940, when Ernst Mayr proposed the "biological" definition of species, or the Biological Species Concept [BSC]:
"Species are groups of interbreeding (or potentially interbreeding) natural populations which are reproductively isolated from other such groups."
Problems....
One cannot apply the BSC definition to a considerable number of asexual populations.
Application of the BSC toward self-pollenating plants is (by its own definition) somewhat limiting.
Moreover, it is often very difficult to apply (practically) anyway. It requires breeding tests to verify the separation, but this is rarely used. Indeed, were breeding tests to be used to define such criterea, natural environments would potentially play an overiding role.
e.g. Do Homo erectus, Homo neanderthalensis and Homo sapiens represent the same or different species? This question is unresolvable using the BSC definition, especially when (as we now suspect) Neanderthals and Homo sapiens may have interbred.
Other Definition of Species include: The Phenetic (or Morphological) Species Concept -Cronquist (1988)- who proposed an alternative to the BSC that he called a "renewed practical species definition", where he very clearly defines species as
"... the smallest groups that are consistently and persistently distinct and distinguishable by ordinary means."
As you can see, determining if two populations are actually different "species"a can be quite a difficult task, especially as the very process of speciation is often a gradual process.
In a more current definition of the term "species", the concept of genetic integration has been coined, and used to differentiate between SOME types of species.
Genetic integration works on the premise that if individuals within a population mate with one another, but not with individuals of other populations, they are considered to be an "independent evolutionary unit", and can safely be called a Species.
But just how do/did New "Species" arise?
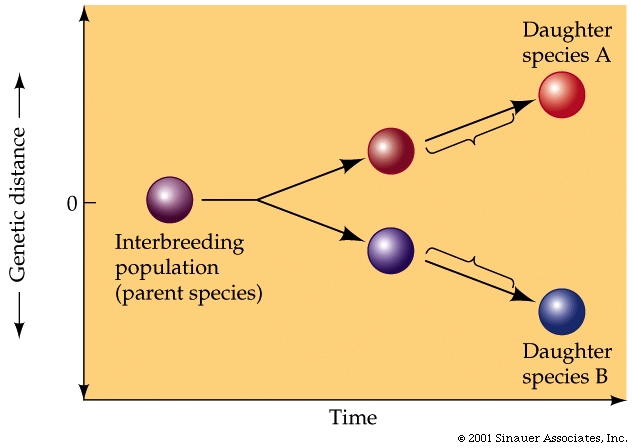
Speciation is the process by which one "species" splits into two.
We have already observed some possible mechanism of a potential speciation....have we not?...in the "black-bellied seed cracking finches of West Africa. But not all such evolutionary changes result in new species. The critical process in the formation of new species is the segregation within the gene pool of the ancestral species into two separate gene pools. Consequently, such speciation is often facilitated by an interruption of gene flow between or among different populations.
Allopatric speciation requires total genetic "reproductive" isolation.....or, when two or more parts of a single population become divided by a geographic barrier, alternatively known as geographic speciation.
Allopatric Speciation is thought to be the dominant form of speciation among most groups of organisms.
Even so, Darwin's seed-eating and bud-eating finches of the Galapagos archipelago demonstrate the importance of geographic isolation as a distinction that (along with population size, perhaps) can enhance the process of speciation.
However, the effectiveness of any barrier -geographical or otherwise- in preventing gene flow depends on the size and mobility of the species in question.
eg. an impenetrable physical barrier for a terrestrial snail may be not be as remarkable a hurdle for a butterfly.
Indirect evidence that most speciation among animals is allopatric is provided by patterns of species distribution.
These findings can lead to a tantalizingly plausible (but ultimately false) "aboslute" inferences......... if that geographic barriers always give rise to speciation....all organisms that are separated by a geographical boundary become species......long-distance romances don't work?
Unfortunately, it is not true...long-distance relationships can work.
Geographic isolation does not necessarily lead to reproductive incompatibility, as evidenced by the European and American sycamores.
Apparently, 20 million years of separation isn't such a long time if neither partner has to change too much.
Consequently, "physical" barriers need not be the only part of the "species" equation.
Anything that begins to separate the gene pool of the ancestral species will suffice.
Speciation, in the absence of any geographical barrier -or the subdivision of a gene pool when members of the daughter species are not geographically separated, but the two species become distinct as a function of inherent genetic polymorphism- is called...
Sympatric speciation.
Any examples spring to mind??
(A) American Sycamores?
(B) West African Finches
(C) Long necked Giant Toitoise in the Galapagos Islands
(D) Birds and Bats
(E) Red winged Blackbirds
Additionally:
Fruit flies have speciated sympatrically in New York State for more than a hundred years. These fruit flies originally courted, mated, and deposited eggs only on hawthorn fruits.
About 150 years ago, large commercial apple orchards were planted in New York. Some fruit flies began to lay their eggs on the apple trees, perhaps by mistake.
Consequently, their offspring sought out apple trees as adults and, therefore, mated with other fruit flies of similar heritage.
Today there are two sympatric species of picture-winged fruit flies in New York.
Peripatric speciation where separation of populations has occurred through some transient acts of nature that have "dispersed a larger poopulation in to far flung, and relatively isolated, pockets -away from the larger population.....
Parapatric speciation separates adjacent population, that have separated as a consequence of reproductive isolation coupled with some degree of migration, again -in the absence of any geographic barrier. Parapatric speciation is quite similar to sympatry speciation, but reflects some migration.
eg. Metal-tolerant and metal-intolerant plants flower at different times, reducing gene flow between the two groups.
Metal-tolerant plants also self-pollinate more frequently.Consequently, reproductive isolation between metal-tolerant and metal-intolerant plants could, therefore, become complete.....potentially giving rise to two new species
eg. Temporal isolation of the three species of Rana Frogs..
Once a barrier to gene flow is established, the resulting daughter populations are then free to diverge genetically -as a consequence of the remaining evolutionary "forces".
In Summary......
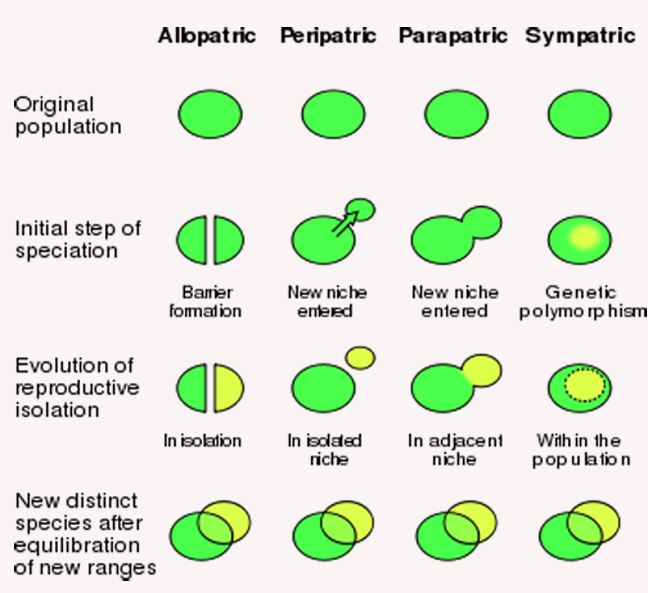
The potential for populations that -for one reason or another- have expanded and diversified their habitats leads to an important evolutionary concept, which is adaptive radiation, which decribes the rapid diversification of organisms as they quickly adapt to newly available resources..... Darwin's finches and kingfishers?
As with all "spec-ial" (speciation) events... the consequences to these separations can be quite different.
Curiously, sometimes, even species that can't reproductivley interact with each other can still share a gene pool ??
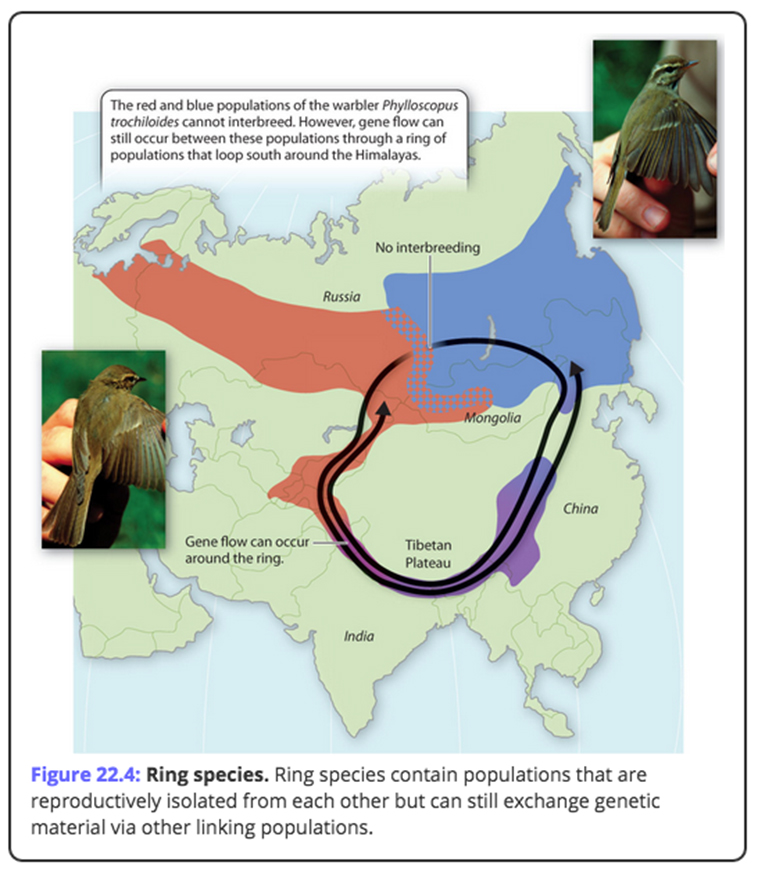
There is no set "degrees of separation" or requisite numbers of generations that define "variations".
Many "variations" of orchids can cross pollenate, while different "species" of cats cannot.

With all its faults, the very foundation stone of the BSC line of thought - the ability to define "species" as a consequence of their reproductive isolation, remains an obvious and powerful "tool" for Biologists who are trying to defining "species" and in determining how species may have come about -at least for the more modern varieties.
BSC also promotes a potential number of mechanisms that may allow for reproductive isolation (i.e. mechanisms that may dictate the rate of accumulation of traits that begin to influence / actually prevent interbreeding between different populations).
Collectively, these mechanisms can help us understand how species are able to form.
Reproductive Isolating Mechanisms
Prezygotic barriers-operate before mating
Five prezygotic reproductive barriers have been described:
Spatial isolation -is the separation by location of inhabitance in the environment.
Temporal isolation -is the separation by differing mating seasons, or times of day.
Mechanical isolation -is the separation by differing shapes of reproductive organs.
Gametic isolation -is the separation by the inability of sperm from one species to fertilize the egg of another species.
Behavioural isolation -is the separation by behaviour, e.g.. diet, lack of recognition of potential mates as mating partners (Agelaius phoeniceus).
Postzygotic barriers -operate after mating.
If individuals of two different "species" still recognize one another and are able to mate, postzygotic reproductive barriers may prevent gene exchange.
There are three postzygotic reproductive barriers have been described, with the major two being:
Low Hybrid viability -the offspring just generally don't do so well.
Hybrid zygote abnormality -which is the failure of a hybrid zygote to develop in to a reproductively viable stage of life.
Hybrid infertility - the inability of a hybrid to reproduce. Hybrid offspring may survive less well than offspring resulting from matings within each species. All the offspring of one sex may be sterile, or all the offspring may be of only one sex. In nearly all cases of hybrid sterility and inviability, it is the sex that is heterozygous for the sex chromosomes that is absent or sterile.
Mules / Hinnys -derived from donkeys and horses- are one of the more readily known examples of this type of barrier, with a female mule only being born every once in a while.
Variation in Speciation Rates.
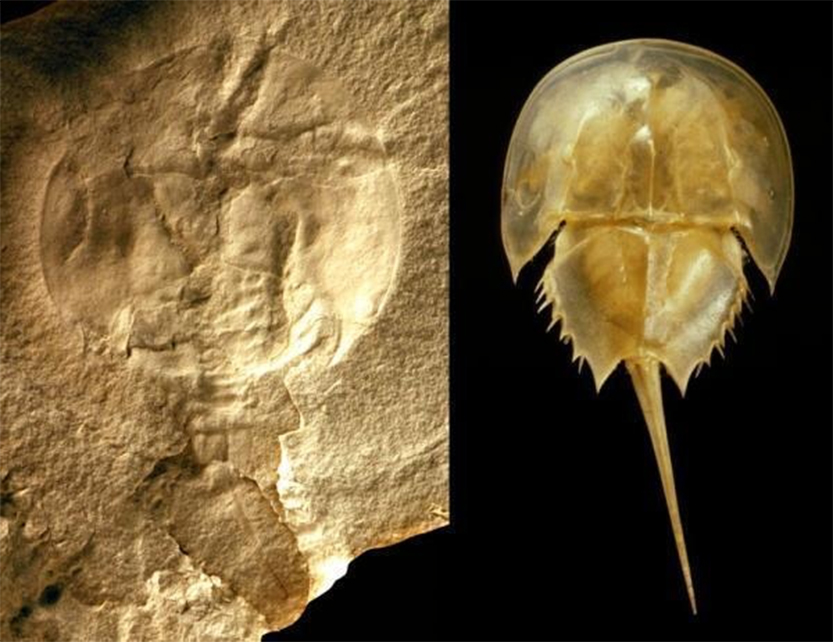
As alluded to previously with the sycamores separated by 20 million years, give or take a few, there are a number of quite distinct factors that effect the rate of speciation. It can be rapid (as in the case of the fruit flies in Hawaii), or it can be relatively slow, as in the case of horse-shoe crabs, where there are only a few of species in existence, but they have survived, relatively unchanged for 445 million years.
Species may differ in relatively few genes.
Species richness: the larger the number of species, the greater will be the number of opportunities for new species to form, even though there may be few actual differences among the different species. Molecular biology techniques allow scientists to measure genetic differences among species.
Studies of Hawaiian fruit flies are demonstrating that many species, may be genetically very similar.
Behaviour may influence speciation rates
Populations of land snails may be separated by barriers as narrow as city streets, which -for the common garden snail- can be quite impressive barriers. Animals with complex behavior may speciate at a high rate because of choices of mates. Indeed, mate selection is probably a major contributor to rapid evolution as a consequence of reproductive isolation between species. Moreover. extremes of sexual selection: complex mating rituals, for example, may differentiate potential partners...parapatric speciation perhaps?Population bottlenecks: the significant alteration in the gene pool, resulting form a "bottleneck" or "founder-effect" change, may result in new adaptive changes (within the population) that result in more rapid speciation.
Environmental changes: may trigger high speciation rates. A climatic change in Africa from warm/wet to more rapid oscillations between warm/wet and cooler/drier conditions may result in a burst of changes.
Evolutionary lineages may also radiate when they acquire a new adaptations that enable them to use the environment in a new way.
Stronger gaping muscles in some species of American blackbirds allow them to turn over objects on the ground where prey may be hiding. With the adaptation of "gaping", blackbirds have come to occupy nearly all habitat types in North and South America.2019nCoV Wuhan China. So, if viruses adapt and evolve, wouldn't that suggest that they ARE alive??

{kind=link}
{kind=link}

About Contact GSU Academics Degrees & Majors |
Admissions Undergraduate Research Dept. of Biology |
Libraries University Library Campus Life Housing |
Athletics Alumni
|